Na Redona és un illot de 1,07 km2 situat just al nord de Cabrera. Lucas Lacasa, de l’Institut de Física Transdisciplinar i Sistemes Complexos de Palma de Mallorca, i Victor M. Eguíluz, del Centre Basc sobre Canvi Climàtic, utilitzen aquest ecosistema en una recerca sobre l’estructura de la participació de les espècies i de les seves funcions en una xarxa ecològica multinivell. Amb Sandra Hervias-Parejo i Mar Cuevas-Blanco com a primeres autores ho expliquen en un article a la revista Nature Communications. L’objectiu és comprendre com les interaccions biòtiques conformen els ecosistemes i influeixen en el seu funcionament, resiliència i biodiversitat. Habitualment, hom tracta per separat les diferents funcions biològiques, però això implica passar per alt l’efecte sinèrgic que hi ha entre elles i no permet captar tots els patrons de participació d’espècies i de funcions. Hervías-Parejo et al. proposen un marc matemàtic que integra diversos tipus d’interaccions biòtiques. L’apliquen a una sèrie de dades recollides recentment a Na Redona, i que abasten 1537 interaccions entre 691 espècies de plantes, animals i fongs. Aquestes interaccions reflecteixen sis tipus de funcions ecològiques: la pol·linització, l’herbivorisme, la dispersió de llavors, la descomposició, la captació de nutrients i la patogenicitat fúngica. Hervías-Parejo et al. troben una estructura no-aleatòries i niuada en la forma que les espècies de plantes participen en diferents funcions. Poden al mateix temps identificar un rang d’espècies i funcions, i trobar que arbustos i descomposició fúngica són actors cabdals de l’ecosistema. El marc d’Hervías-Parejo et al. és dual en el sentit que compagina una perspectiva d’espècie i una perspectiva de funció tant en la quantificació de la complexitat d’un ecosistema com en la determinació del seu funcionament i diversitat.
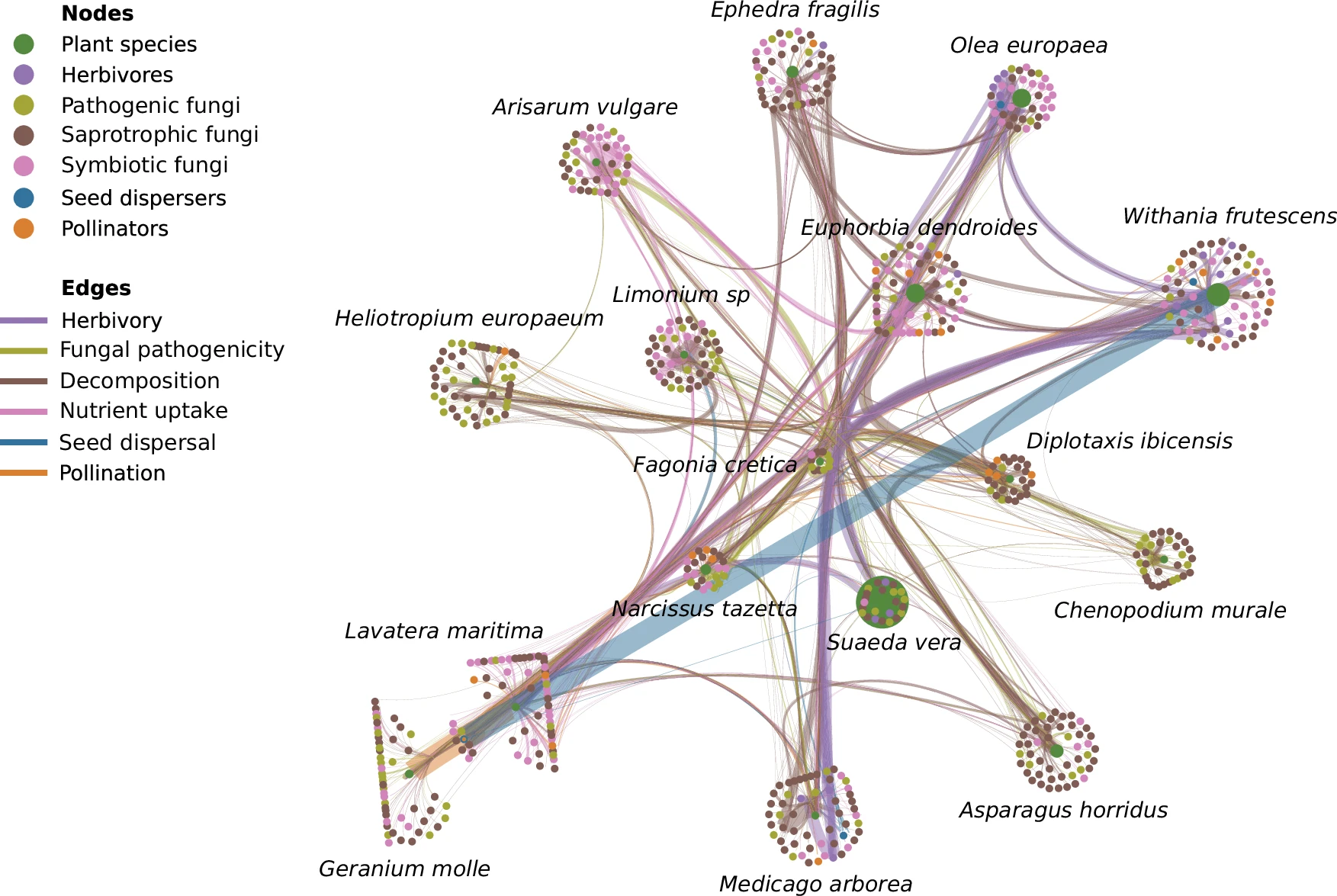
Hervías-Parejo et al. han elaborat un tensor empíric sobre la funció recurs-consumidor entre 691 espècies que habiten l’illa de Na Redona, a l’arxipèlag de Cabrera. S’interessen en sis funcions: pol·linització, herbivoria, dispersió de llavors, descomposició de matèria vegetal, captació de nutrients per micorrizes i patogenicitat fúngica.
La recerca a Na Redona
Anna Traveset (IMEDEA), Ruben Heleno (CFE), Sandra Hervías-Parejo (IMEDEA, CFE) i Susana Rodríguez-Echeverría (CFE) conceberen i dissenyaren la recerca per construir la xarxa multinivell empírica i recollir dades de camp. Hervías-Parejo liderà el tractament de dades posterior a la identificació de tots els tàxons, i construí la xarxa multinivell. Rodríguez-Echevarría classificà els fongs en els tres grups funcionals de sapròfits, simbionts i patògens. Lucas Lacasa (IFISC) i Victor M. Eguíluz (BC3) conceptualitzaren el marc i guiaren la modelització matemàtica. Mar Cuevas-Blanco (IFISC) contribuí a la modelització matemàtica, realitzà l’anàlisi de xarxa i de dades, les simulacions i generà la majoria de gràfics. Lacasa, Eguíluz i Carlos J. Melián (IFISC) contribuïren a l’anàlisi de dades. Hervías-Parejo, Cuevas-Blanco, Lacasa, Traveset, Isabel Donoso (IMEDEA, CFE), Melián i Eguíluz discutiren els resultats. Lacasa, Cuevas-Blanco i Eguíluz escrigueren el primer esborrany de l’article. Lacasa, Eguíluz, Hervías-Parejo, Cuevas-Blanco, Traveset i Donoso escrigueren i revisaren l’esborrany definitiu. Aquest fou tramès a Nature Communications el 3 d’agost, que l’acceptà el 27 de setembre i publicà el 23 d’octubre.
El protocol d’estudi fou revisat i aprovat pel Comitè Ètic del Centre Superior d’Investigacions Científiques (CSIC) amb el codi CGL2017-88122-P, PID2020-114324GB-C21.
El treball de camp es realitza a Na Redona (39° 10′ 5′′ N, 2° 58′ 35′′ E), un illot d’11 hectàrees i una altitud màxima de 56 metres que fa part del Parc Nacional de l’Arxipèlag de Cabrera. L’hàbitat primari és un matoll mediterrani amb una relativa riquesa en termes d’espècies de plantes, unes 108. El treball de camp inclogué el mostreig i captura de sargantanes i ocells, amb els permisos de les autoritats del Parc Nacional (SEN 184/18, SEN 074/19, SEN 687/2020). Ruben Heleno (CFE Coimbra) capturava els ocells sota la llicència 221/2019 de l’Instituto da Conservação da Natureza e das Florestas. Manuel Nogales (IPNA-CSIC) capturava les sargantanes amb la llicència CAP-169.4-20 de la Dirección General de Agricultura, Ganadería y Alimentación.
El treball de camp es duia a terme en dues estacions diferents, en el pic de la floració (abril i maig) i en el pic de la fructificació (octubre i novembre). Durant cinc dies consecutius un equip de cinc persones visitava l’illa per mostrejar els diferents tipus d’interacció entre plantes i pol·linitzadors, herbívors, dispersadors de llavors, fongs colonitzadors d’arrels (micorrizes), fongs saprofítics, fongs simbiòtics i fongs patogènics. En cada estació de mostreig s’establien sis transsectes (cadascun de 100 metres de llarg i 10 metres d’amplada, i separats cadascun de l’altre per un mínim de 100 metres) per tal de cobrir els principals microhàbitats i tot el gradient altitudinal de l’illa. En cada transsecte s’estimava l’abundància de plantes (nombre d’individus) i la coberta vegetal (en metres quadrats) de cada espècie.
Les interaccions planta-fong se seguien a través de la recol·lecció de les arrels de cinc individus de cada espècie al llarg dels transsectes. Les arrels netejades en sec eren preservades en silica gel fins a processar-les per extreure’n l’ADN. Sobre l’extracte d’ADN es feina una amplificació per PCR amb els encebadors fúngics ITS1f (5′-CTTGGTCATTTAGAGGAAGTAA-3′) i ITS2 (5′-GCTGCGTTCTTCATCGATGC-3′). Els amplicons resultants eren netejats, quantificats i seqüenciats en una plataforma Illumina a GENYO de la Universidad de Granada. Les lectures eren assignades a variants ASV.
Les interaccions planta-pol·linitzador eren estudiades amb censos de 10 minuts de durada en diferents moments del dia. En aquest cens es feia una observació directa de cada planta individual. Hom anotava tot animal (insecte, sargantana o ocell) que contactés amb les parts reproductives les flors. Els insectes eren fotografiats i identificats posteriorment a IMEDEA. Es capturaren 30 sargantanes a llaç i 26 aus terrestres amb xarxes. De les sargantanes es prenia una mostra del morro per trobar-hi grans de pol·len al microscopi a IMEDEA.
Les interaccions planta-herbívor eren avaluades en cinc individus de cada espècie de planta. S’examinaven branques aïllades i es registraven tots els artròpodes trobats alimentant-se de teixits vegetals.
Les interaccions planta-dispersador de llavors es basaven en la recol·lecció d’excrements i pellets de gavines (236), d’excrements de passerins (21) i sargantanes (375) i material de formiguer (4). En aquestes mostres s’identificaven restes de plantes a l’estereomicroscopi a l’IMEDEA.
Els autors agraeixen a Xavier Canyelles la identificació de tots els insectes; a José Antonio Morillo per l’anàlisi bioinformàtica de les interaccions planta-fong; a Aarón González-Castro i Juan Pedro González-Varo per l’ajut en el treball de camp; a Araceli Guillem Bosch per l’ajut en el processament de mostres de pol·len al laboratori; a Miguel A. Muñoz pels seus comentaris sobre l’avaluació del pes de cada espècie en xarxes mutualistes. Hervías-Parejo rep finançament del Govern Balear pel projecte PD/004/2019. D’altres autors reben finançament de l’agència espanyola de recerca i de la fundació portuguesa de ciència i tecnologia.
Ecosistemes: espècies i funcions
Un ecosistema és biocenosi i biòtop. La biocenosi és el conjunt d’organismes vius que en formen part. El biòtop és el medi físic on es relacionen aquests organismes. Podem definir-lo també com un conjunt d’espècies que interactuen entre elles. De fet són aquestes interaccions les que determinen el nínxol ecològic (entès a la manera de Charles Elton) que ocupa cada espècie. El nínxol eltonià és multidimensional, ja que múltiples són els papers ecològics o funcions que descriuen les interaccions. Com a exemples de funcions ecològiques poden enumerar la pol·linització, la dispersió de llavors, la captació de nutrients o la descomposició.
La descripció d’un ecosistema sol posar al centre les espècies que hi coexisteixen. Seguidament uneix els diferents nodes (les diferents espècies) d’acord amb les interaccions que s’hi estableixen. El més habitual és que la descripció contempli un tipus concret d’interacció. Així, per exemple, la xarxa tròfica reflecteix qui consum a qui; les xarxes mutualístiques reflecteixen les interaccions mútues; etc. La ciència de xarxes ha utilitzat aquesta aproximació per descriure cascades d’interaccions d’espècies o per deduir la importància de l’estructura ecològica sobre la biodiversitat.
Quan hom contempla al mateix temps diversos tipus d’interacció pot recórrer a la teoria de xarxa multinivell. Cada nivell es correspon a un tipus d’interacció. A multinivell contemplem la multifuncionalitat de l’ecosistema.
Ara bé, aquesta xarxa multinivell requereix observacions empíriques molt fines sobre les interaccions entre espècies. A més, aquestes observacions solen requerir mètodes diferents, difícils d’integrar en l’anàlisi posterior. Finalment, la mètrica de la xarxa multinivell no es troba dan desenvolupada com la de la xarxa convencional.
El funcionament dels ecosistemes es pot descriure alternativament a través de funcions, o del concepte d’espècie-funció. En aquest cas el node ja no és l’espècie com a tal, sinó l’espècie-funció.
Hervías-Parejo et al. parteixen del paradigma consumidor-recurs. En aquest paradigma les plantes, com a productors primaris, són considerades ‘recurs’. Animals i fongs, per la seva banda, són considerats consumidors.
La tria de Na Redona com a subjecte experimental no és casual. L’illot representa una comunitat relativament simple. I diem relativament, perquè amb la seva metodologia de mostreig estandarditzat, Hervías-Parejo et al. han identificat 1537 interaccions ponderades entre plantes, animals i fongs. Les interaccions es poden classificar en sis funcions ecològiques: 1) pol·linització; 2) herbivorisme; 3) dispersió de llavors; 4) descomposició; 5) captació de nutrients i 6) patogenicitat fúngica.
El primer pas és transformar aquesta base de dades d’interaccions en un tensor de la funció recurs-consumidor. Aquest tensor RCF es pot entendre com una xarxa ponderada multifuncional. Del tensor es deriva un índex de consumidor i una matriu recurs-funció. Amb una anàlisi d’extinció simulada s’avalua la importància d’espècies i de funcions en l’estructura de l’ecosistema.
La construcció del tensor RCF
Resulta difícil captar la multidimensionalitat d’un ecosistema. Hervías-Parejo et al. parteixen de dades que inclouen l’observació directa de 16 espècies de plantes, de 629 variants de seqüència d’amplicó de fongs i de 46 espècies animals. Cada dimensió representa una de les següents funcions: 1) pol·linització; 2) herbivorisme; 3) dispersió de llavors; 4) descomposició fúngica; 5) captació fúngica de nutrients; i 6) patogenicitat fúngica.
L’element fonamental és com un recurs (és a dir una espècie de planta) i interactua amb un consumidor (animal o fúngic) x a través d’una funció α. Sobre això es basteix un tensor de rang 3, o cada element especifica la probabilitat observada de cada esdeveniment. El recurs i i el consumidor x són els índexs covariants del tensor, i la funció α és l’índex contravariant.
Es tracta efectivament d’una visió fitocèntrica de les funcions ecològiques. Al capdavall el punt de partida és descriure en quina mesura l’espècie de planta i interactua a través d’α amb el consumidor x: el valor pot anar de 0 (mai) a 1 (sempre).
Una xarxa ecològica multinivell
Podem entendre que els nodes de la xarxa són les diferents espècies. Cadascuna de les 16 espècies de plantes es relaciona amb els 675 consumidors a través de sis relacions. Així la xarxa consta de sis nivells o capes.
El caràcter niat de la matriu recurs-funció
Els diferents recursos (les 16 espècies de plantes) i les diferents funcions (les 6 funcions ecològiques) es poden descriure en una matriu. Per fer-ho cal integrar els índexs de consumidors del tensor RCF, per així aconseguir la probabilitat de participació de cada recurs en cadascuna de les sis funcions ecològiques. La matriu resultant és un tensor de rang 2. Hervías-Parejo et al. en destaquen l’estructura niada. A aquesta estructura hi contribueixen tant les espècies de plantes com les funcions ecològiques, ja que totes dues dimensions són heterogènies.
Per exemple, l’orval (Withania frutescens) i la malva roquera (Lavatera marítima) participen en les sis funcions ecològiques amb probabilitats superiors a 0,7. Són, doncs, plantes generalistes en termes de funció.
Per contra, el blet de mur (Chenopodium murale) i la cua d’escorpí (Heliotropium europaeum) participen en menys funcions. Podem dir que són especialistes en termes funcionals.
Les espècies clau
Les espècies de plantes que són generalistes en termes de funcions actuen com a connectors funcionals. Hervías-Parejo et al. construeixen xarxes funció-funció per trobar-hi les espècies de plantes que hi participen. Per fer-ho projecte la matriu recurs-funció en la classe de funció. En resulta una xarxa amb 6 nodes (un per cada funció ecològica), amb un rang d’1.
Withania frutescens i Lavatera marítima apareixen com a espècies multifuncionalment cabdals. Hervías-Parejo et al. comproven que aquest caràcter cabdal no correlaciona amb l’abundància relativa d’aquestes espècies ni tampoc amb el grau de cobertura vegetal.
Les funcions cabdals
Alternativament podem considerar com les diferents funcions ecològiques connecten les diverses espècies de plantes. Ara la matriu recurs-funció és projectada en la classe de plantes resultant-hi una xarxa efectiva de 16 nodes.
La funció que més destaca és la descomposició. Al capdavall és el destí comú de tota la matèria viva.
La voluntat de superar dos reptes
El primer repte que es proposaven Hervías-Parejo et al. era la incorporació de diferents funcions ecològiques en una xarxa multinivell. Ho fan a través d’un tensor de rang 3.
Quan redueixen el tensor RCF a una matriu recurs-funció posen de manifest un patró niat en la forma que recursos i funcions participen entre ells. Es fa possible mesurar quines espècies de plantes són cabdals en l’estructura de l’ecosistema. Curiosament són plantes llenyoses com Withania frutescens i Lavatera marítima les que tenen una major puntuació en termes de multifuncionalitat. Les plantes llenyoses tenen un cicle vital més llarg que les plantes herbàcies.
Pel que fa a les funcions cabdals destaca la descomposició. La descomposició és un tipus d’interacció planta-fong de caràcter saprofític. Cal recordar que a Na Redona hi ha poques plantes amb fruits carnosos, de forma que la dispersió de llavors no se sol fer amb una interacció biòtica.
El model d’Hervías-Parejo et al. no té en compte les interaccions planta-planta o les interaccions consumidor-consumidor. Per fer-ho, caldria incorporar més índexs al tensor, o bé considerar totes les possibles interaccions entre espècies. Hervías-Parejo et al. s’orienten més cap a la segona possibilitat, però són conscients que això requeriria superar les observacions purament fitocèntriques, incorporant dades com ara la trajectòria dels insectes pol·linitzadors.
Hervías-Parejo et al. consideren que el seu marc té aplicacions més enllà de l’ecologia cap a altres tipus de sistemes complexos. Per exemple, el mapa recurs-funció pot traslladar-se als mapes de genotip-fenotip de la genètica, als gràfics d’interacció econòmica entre països, etc.
Lligams:
– On the structure of species-function participation in multilayer ecological networks. Sandra Hervías-Parejo, Mar Cuevas-Blanco, Lucas Lacasa, Anna Traveset, Isabel Donoso, Ruben Heleno, Manuel Nogales, Susana Rodríguez-Echeverría, Carlos J. Melián, Victor M. Eguíluz. Nature Communications 15: 8910 (2024).
– On the structure of species-function participation in multilayer ecological networks. Mar Cuevas Blanco. Github.
