Raúl Méndez i Xavier Salvatella són investigadors de la unitat de Mecanismes de Malalties de l’Institut de Recerca Biomèdica de Barcelona. L’autisme és una de les patologies en les quals són interessats. L’entenen com una condició del neurodesenvolupament que afecta l’adquisició d’habilitats de comunicació i d’interacció social. La genètica ha permès explicar aproximadament un 20% dels casos d’espectre autista a través de mutacions específiques. Cal pensar que en el 80% restant, l’anomenat ‘autisme idiopàtic’, la base neurològica és més complexa. El grup de Méndez i Salvatella va descriure en el seu moment que una disminució de la inclusió d’un microexó de 24 nucleòtids en l’ARNm que codifica per a la proteïna neuronal CPBE4 té un vincle amb el trastorn d’espectre autista idiopàtic. Ara, en un article a Nature, amb Carla Garcia-Cabau de primera autora, aprofundeixen en com aquest fenomen de splicing incorrecta condueix a una agregació de la proteïna CPEB4. La transcripció de gens codificadors de proteïna genera un ARN immadur que, entre d’altres coses, patirà un splicing mitjançant el qual seran seleccionats les parts del gen que formaran part finalment de l’ARN missatger madur. Els exons són les regions del gen que poden aparèixer en l’ARNm, mentre que els introns són les regions que no ho fan mai. Ara bé, sovint els gens poden ser objecte d’un splicing alternatiu, de manera que no tots els exons potencials apareixeran en totes les formes finals d’ARNm d’aquell gen. En proteïnes neuronals és habitual la inclusió mitjançant aquest splicing alternatiu de microexons. Els microexons específics de neurones són objecte d’una recerca intensa, ja que hom sospita que poden jugar un rol en el neurodesenvolupament. El grup de Méndez i Salvatella, com hem dit, havia mostrat ja que la inclusió d’un microexó de 24 nucleòtids específic de neurones en la proteïna CPBE4 juga un paper potencial en l’autisme idiopàtic. La proteïna CPBE4 és una proteïna d’unió a ARN que regula la traducció d’ARNm induint-hi canvis en la longitud de la cua de poliadenina (poliA). Entre els ARNm que regula hi haurien els de gens altrament vinculats a l’espectre autista idiopàtic. Ara Garcia-Cabau et al. mostren en un model de ratolí que la proteïna neuronal CPEB4 forma condensats que es dissolen després de la despolarització. Aquesta transició s’associa amb un interruptor que passa de la repressió a l’activació traduccional. Interaccions heterotípiques entre el microexó i un clúster de residus d’histidina evita l’agregació irreversible de CPEB4 a través de la competència amb les interaccions homotípiques entre clústers d’histidina. Així doncs, el microexó en qüestió és clau perquè CPEB4 mantingui la reversibilitat entre les formes agregades i desagregades i així participi en la regulació de l’expressió de tot un ventall de gens en resposta a l’estimulació neuronal.
Garcia-Cabau et al. han estudiat en un model de ratolí la condensació de la proteïna CPEB4 en neurones.
Les bases neuronals de l’espectre autista
Diversos estudis han posat de manifest que en individus amb trastorn de l’espectre autista hi ha una desregulació dels processos d’splicing alternatiu que coordinen la inclusió de microexons específics de neurones en els ARNm que es tradueixen en proteïnes neuronals.
Un exemple d’aquests microexons és el microexó 4 (me 4) del gen CPEB4. Aquest microexó consisteix en una seqüència de 24 nucleòtids. El grup de Méndez i Salvatella ha mostrat que en persones amb autisme idiopàtic o amb esquizofrènia hi ha una menor inclusió de me 4 en la proteïna CPEB4.
La proteïna CPEB4 pertany a la família de proteïnes CPEB. Són proteïnes d’unió a ARN que s’uneixen als transcrits d’ARN a través de la seqüència CPE. La seqüència CPE és l’element citoplasmàtic de poliadenilació, i es troba en la regió 3’ dels transcrits que no es tradueix a proteïna. Les proteïnes CPEB, en unir-se a la seqüència CPE, intervenen en la poliadenilació o deadenilació del transcrit d’ARN. La cua de poliadenina (poliA) és un dels trets d’ARNm madur i regula la traducció d’aquest ARNm a través de l’ensamblatge dels complexos ribonucleoproteics responsables de la traducció.
Les proteïnes CPEBs participen en el transport i localització d’ARNm, i juguen un paper important en processos com la divisió cel·lular, el desenvolupament neural, l’aprenentatge i la memòria.
Les proteïnes CPEBs es poden classificar en dues subfamílies segons els motius de reconeixement d’ARNm, d’interacció amb altres proteïnes i de regulació. El domini d’unió a ARN es troba en la regió carboxi-terminal de la proteïna CPEB i és globular, mentre que els dominis de la regió amino-terminal (NTDs) són més desordenats. La fosforilació de múltiples residus de serina i treonina en els NTDs de CPEB4 per part d’ERK2 i CDK1 activa la proteïna CPEB4 en dissoldre els condensats repressors de la traducció.
La CPEB4 neuronal (nCPEB4) s’uneix a transcrits d’un ample ventall de gens, un bon nombre dels quals s’han vist associats al trastorn d’espectre autista (ASD).
Un model de ratolí que expressa desproporcionadament la isoforma nCPEB4 que manca del me4 (nCPEB4Δ4) presenta els defectes de poliadenilació i d’expressió de proteïna observats en ASD, i també expressa fenotips neuroanatòmics, electrofisiològics i comportamentuals típics de l’ASD.
La isoforma nCPEB4Δ4 es comporta com una variant negativa dominant en neurones, però en canvi és activa en teixits no-neuronals.
Les neurones es caracteritzen per ser, en termes generals, cèl·lules que no segueixen ja el cicle cel·lular mitòtic. Una altra característica diferencial té a veure amb la despolarització de membrana ja que, de fet, l’impuls nerviós és bàsicament una ona de despolarització neuronal. La despolarització neuronal implica canvis en el pH intracel·lular i això afecta el grau de condensació de la proteïna nCPEB4. La proteïna nCPEB4Δ4 tendeix a formar agregats irreversibles. El rol de me4 sembla consistir en assegurar el caràcter reversible dels condensats de nCPEB4.
La regulació de la condensació nCPEB4
La proteïna CPEB4 s’expressa en la majoria de teixits, però ho fa de manera més abundant en el sistema nerviós. El microexó 4 (me4) únicament apareix en la forma neuronal de CPEB4 (nCPEB4). La raó d’això és que únicament en neurones s’expressa SRRM4, regulador de l’splicing requerit per a la inclusió de microexons com me4.
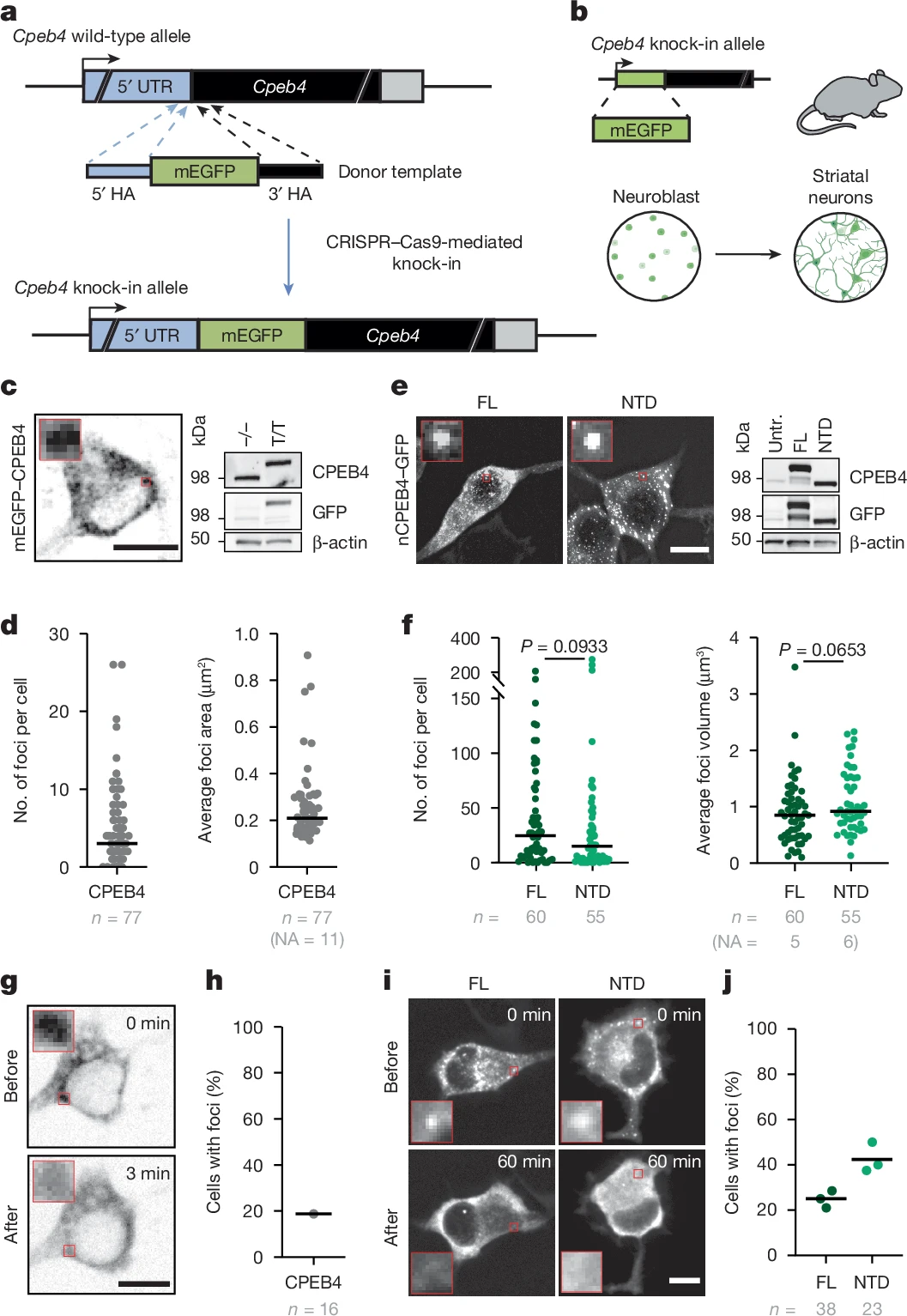
Garcia-Cabau et al. han generat un model de ratolí knock-in per a mEGFP–CPEB4. D’aquests ratolins obtenen neurones estriatals primàries. En aquestes neurones CPEB4 té una distribució citoplàsmica granular que recorda els condensats de sobreexpressió de CPEB4 de cèl·lules no-neuronals.
D’aquest model de ratolí knock-in per a mEGFP–CPEB4 també obtenen cèl·lules neuronals N2a. Aquestes són neurones en procés de maduració. Quan marquen la nCPEB4 amb proteïna fluorescent verda (GFP), troben la formació de condensats.
Garcia-Cabau et al. han investigat com afecta la despolarització ex vivo de neurones amb KCl o NMDA a les propietats dels focus o condensats de nCPEB4. El KCl despolaritza les cèl·lules N2a i dissol els condensats de nCPEB4 (tant de la proteïna plena com del domini amino-terminal). El NMDA no despolaritza les cèl·lules N2a.
García-Cabau et al. han estudiat in vitro el comportament del domini N-terminal (NTD) de nCPEB4. La calor condueix a la condensació de NTD.
Mitjançant espectrometria de masses, García-Cabau et al. investiguen si modificacions post-traduccionals poden provocar la dissolució de condensats de nCPEB4 després de la despolarització de cèl·lules N2a. Els resultats indiquen que la fosforilació de nCPEB4 no sembla gaire rellevant en la dissolució dels condensats de nCPEB4. Tampoc no sembla gaire rellevant la mobilització de calci, ja que la ionomicina no altera els condensats per ella mateixa.
Amb el pigment SNARF-5F, Garcia-Cabau et al. estimen el pH intracel·lular de N2a. La despolarització en aquestes cèl·lules fa baixar el pH en 0,08 unitats durant uns 10 minuts. Després el pH puja fins a 0,3 unitats. Aquests canvis de pH són prou rellevants com per afectar els cadenes laterals dels residus d’histidina de nCPEB4(NTD).
Constructes que substitueixen histidines amb serines en la nCPEB4(NTD) afecten la propensió a la condensació. Sembla doncs que el caràcter aromàtic dels residus d’histidina és crucial en la propensió a la condensació de nCPEB4(NTD), possiblement per la interacció entre els anells benzènics i cadenes laterals carregades positivament d’altres residus, com ara aspartats i glutamats, o bé arginines.
El microexó 4 prevé l’agregació de CPEB4
Garcia-Cabau et al. empren l’espectroscopia RMN en solució per identificar els residus responsables de la condensació de nCPEB4(NTD). En condicions de pH elevat i temperatura moderada, nCPEB4(NTD) forma multímers amb un diàmetre hidrodinàmic d’uns 55 nm. Aquesta multimerització i vitro es deuria a interaccions entre residus d’histidina i d’arginina. Això inclou interaccions entre els residus 229-252 de HClust amb els residus 403-410 de me4. La deleció de HClust i/o la de me4 condueix a una disminució de la propensió de nCPEB4(NTD) a la multimerització i condensació. Això no passa tan sols in vitro sinó també en les cèl·lules N2a.
La comparació entre nCPEB4 (proteïna nadiua) i nCPEBΔ4 (proteïna amb deleció de me4) mostra que totes dues tenen interaccions semblants amb altres proteïnes i s’uneixen totes dues de manera similar a ARN.
Els condensats de nCPEB4 estabilitzats per me4 actuen com a repressors de la traducció d’ARN a proteïnes. En el 49% de cèl·lules N2a que sobreexpressen nCPEB4 els condensats d’aquesta proteïna es dissolen després de la despolarització. En canvi, si sobreexpressen nCPEB4Δ4 la proporció cau al 17%. D’alguna manera, doncs, me4 augmenta la reversibilitat de la condensació de nCPEB4. Val a dir que en aquesta reversibilitat no participen modificacions post-traduccional de nCPEB4.
En experiments in vitro, els condensats de nCPEB4(NTD) es dissolen quan baixa la temperatura. En canvi, els condensats de nCPEB4Δ4(NTD) la dissolució es tan sols parcial. La raó d’aquesta diferència es trobaria en la transició del condensats de nCPEB4Δ4(NTD) a agregats. Aquesta agregació no varia encara que hi hagi presència d’ARN.
L’ortòleg de nCPEB4 en la mosca de la fruita (Drosophila melanogaster) és Orb2. El nucli amiloide de Orb2 ha estat caracteritzat per microscòpia electrònica criogènica. Garcia-Cabau et al. utilitzen aquesta estructura per assumir que HClust és responsable de l’agregació in vitro de nCPEB4Δ4(NTD), i que me4 actua com a inhibidor d’aquesta agregació.
En el 78% de cèl·lules N2a que expressen nCPEB4Δ4ΔHC la despolarització condueix a la dissolució dels condensats d’aquesta proteïna. Garcia-Cabau et al. dedueixen que me4 estabilitza cinèticament els condensats de nCPEB4 repressors de la traducció impedint-ne l’agregació: així, quan hi ha la despolarització neuronal, els condensats es dissolen més fàcilment i s’esvaeix la repressió de la traducció.
L’efecte dominant negatiu de l’absència de me4
Quan neurones coexpressen nCPEB4 i nCPEB4Δ4, aquesta darrera té un efecte dominant negatiu sobre la traducció d’ARNm. Les dues variants co-localitzen i tenen dinàmiques i coeficients de partició similars. N’hi ha prou amb una petita porció de nCPEB4Δ4 perquè els condensats agreguin de manera irreversible.
Els ratolins TgCPEB4Δ4 són un model de ratolí del trastorn d’espectre autista. Garcia-Cabau et al. han comparat la conformació in vivo de CPEB4 en cervells de ratolins de 6 mesos de TgCPEB4Δ4 i controls. Una electroforesi en gel d’agarosa amb un detergent semidesnaturalitzant (SDD-AGE) posa de manifest la major presència d’agregats de CPEB4 en el cervell de ratolins TgCPEB4Δ4 respecte de ratolins control. Aquests agregats són resistents a la proteïnasa K. També són capaços d’induir l’agregació in vitro de proteïna soluble CPEB4 de ratolins control. N’hi ha prou, doncs, amb un petit canvi negatiu en el grau d’inclusió de me4 en CPEB4 per promoure l’agregació d’aquesta proteïna i per tant una repressió de la traducció de gens que no es reverteix amb la polarització. En talls ultrafins de neurones estriatals de ratolins TgCPEB4Δ4 hi ha una major presència de condensats de CPEB4 amb signes d’agregació.
Els residus d’arginina de me4 inhibirien l’agregació a través de l’estabilització cinètica dels condensats de nCPEB4, en desplaçar l’equilibri entre les interaccions homotípiques de HClust amb interaccions heterotípiques HClust-me4. Garcia-Cabau et al. han dissenyat un pèptid sintètic que conté dues repeticions de la seqüència de me4. El pèptid és capaç d’entrar en els condensats de CPEB4, d’augmentar-ne l’estabilitat termodinàmica i de disminuir-hi l’agregació.
Un pèptid com a fàrmac contra l’autisme?
La seqüència del pèptid en qüestió és Ac-ARTYGRRR-(GS)3-ARTYGRR-NH2. La seqüència ARTYGRRR és la que deriva del microexó 4 (me4). És una seqüència rica en arginines, i són aquestes arginines les que augmenten l’estabilitat cinètica de la proteïna CPEB4 condensada. Les arginines de me4 interactuen amb un clúster de residus d’histidina (HClust) que es troben en el centre del NTD de la proteïna CPEB4. Podem dir que hi ha una interacció heterotípica entre me4 i HClust, que afavoreix que els condensats de CPEB4 siguin reversibles, dissolent-se quan la neurona experimenta una despolarització. La dissolució dels condensats de CPEB4 permet que siguin traduïts tot un ventall d’ARNm. Aquests ARNm corresponen a gens implicats en el neurodesenvolupament, la repressió dels quals afavoreix símptomes vinculats al trastorn de l’espectre autista.
En absència de me4, les interaccions homotípiques entre HClust condueixen a una agregació irreversible de CPEB4. Llavors els agregats de CPEB4 mantenen una repressió traduccional que ja no és suspesa en moments de despolarització neuronal.
Garcia-Cabau et al. parlen d’un tamponament heterotípic com un mecanisme de preservació del caràcter dinàmic d’un condensat biomolecular. Sense aquest tamponament els condensats degeneren en agregació, i és ben sabut que l’agregació és al darrera de tota una sèrie de proteïnopaties.
En les neurones l’existència d’un splicing alternatiu fa que hi hagi una coexistència de nCPEB4 i nCPEB4Δ4. Aquesta segona isoforma té un efecte negatiu-dominant de manera que n’hi ha prou amb una petita disminució en el grau d’inclusió del microexó 4 (me4) com perquè augmenti amb escreix la repressió que els condensats de CPEB4 fan sobre la poliadenilació de transcrits de gens associats al trastorn de l’espectre autista. Cal que me4 sigui present al menys en un 60% de les proteïnes d’un condensat de CPEB4 perquè aquest mantingui el caràcter líquid que fa que es dissolgui arran dels canvis de pH induïts per la despolarització.
Garcia-Cabau et al. creuen que una baixa inclusió de me4 és un factor patogènic en el trastorn de l’espectre autista a través de l’agregació de CPEB4 i la consegüent baixa expressió de gens vinculats a la protecció davant d’aquest trastorn. En individus amb trastorn d’espectre autista vora el 45% de nCPEB4 és nCPEB4Δ4. Garcia-Cabau et al. consideren necessari aprofundir en el model de ratolí TgCPEB4Δ4 sobre com evoluciona aquest splicing alternatiu durant el desenvolupament embrionari.
Garcia-Cabau et al. també remarquen la rellevància del pH intracel·lular en la dissolució dels condensats de nCPEB4 arran de la despolarització neuronal. Queda per veure si això també passa en altres proteïnes de la mateixa família, com CPEB2 i CPEB3. Pel que fa a la base molecular de l’exclusió de me4, l’atenció hauria de posar-se en el factor de splicing SRRM4, el qual també intervé en la regulació dels microexons de eIF4G.
Un pèptid que incorpora la seqüència me4 podria ser un trans-restaurador de l’activitat normal de CPEB4, i constituir doncs la inspiració per a possibles tractaments del trastorn d’espectre autista.
Lligams:
– Mis-splicing of a neuronal microexon promotes CPEB4 aggregation in ASD. Carla Garcia-Cabau, Anna Bartomeu, Giulio Tesei, Kai Chit Cheung, Julia Pose-Utrilla, Sara Picó, Andreea Balaceanu, Berta Duran-Arqué, Marcos Fernández-Alfara, Judit Martín, Cesare De Pace, Lorena Ruiz-Pérez, Jesús García, Giuseppe Battaglia, José J. Lucas, Rubén Hervás, Kresten Lindorff-Larsen, Raúl Méndez, Xavier Salvatella. Nature (2024).
– Avenç clau en autisme: descobert el paper crucial dels condensats de la proteïna CPEB4. Notícia de l’IRB Barcelona
