En una ocasió anterior (14/2017) ens hem referit a la recerca que fa Jean-François Briand Laboratòri de Materiaus, Polimèrs, Interfàcia i Environament Marítim (MAPIEM) de l’Universitat de Tolon (La Gàrdia) sobre la formació de biofilms en substrats artificials submergits. Aurélie Portas va defensar l’any passat una tesi doctoral elaborada durant quatre anys al MAPIEM, centrada especialment sobre l’impacte negatiu de les incrustacions d’organismes biològics sobre les instal·lacions marines d’energies renovables (eòlica, solar, maremotriu). Un aspecte d’aquesta tesi doctoral és la influència de la hidrodinàmica en la biocolonització de superfícies per part de microorganismes procariòtics i eucariòtics. En un article a Environmental Microbiome, Portas et al. analitzen l’impacte del moviment de l’aigua sobre la dinàmica d’aquestes comunitats microbianes, de la seva producció metabòlica, així com l’impacte sobre la incrustació de microorganismes i del sosteniment de macroorganismes incrusadors i de la fauna vàgil que s’hi associa. Ho han fet en un ambient litoral semi-controlat a la Badia de Tolon. Un dispositiu de camp simulava un gradient de tensió de tall sobre una superfície de PVC. Les comunitats microbianes eren estudiades a través de metabarcoding, cromatografia (LC-MS), tests bioquímics i microscòpia al llarg d’un mes en cadascuna de les quatre estacions. Portas et al. troben que la tensió tallant és el factor primari sobre la biomassa, producció de substàncies polimèriques extracel·lulars i l’estructura i composició de la comunitat microbiana. La transició d’una condició estàtica a una de dinàmica altera la beta-diversitat procariòtica i eucariòtica molt més que no pas els canvis en la intensitat de la tensió de tall o els canvis estacionals de paràmetres físico-químics. En superfícies estàtiques hi predominen artròpodes i nematodes que s’alimenten de microbis. En superfícies en moviment hi ha més aviat organismes sèssils i protistes heterotròfiques, que aprofiten els biofilms com a estructura de protecció per créixer-hi. Si augment el moviment i, en conseqüència, la tensió tallant, llavors hi ha una disminució de la biomassa però alhora hi ha una resposta de protecció en forma d’una major generació d’exopolisacàrids i d’altres biopolímers extracel·lulars.
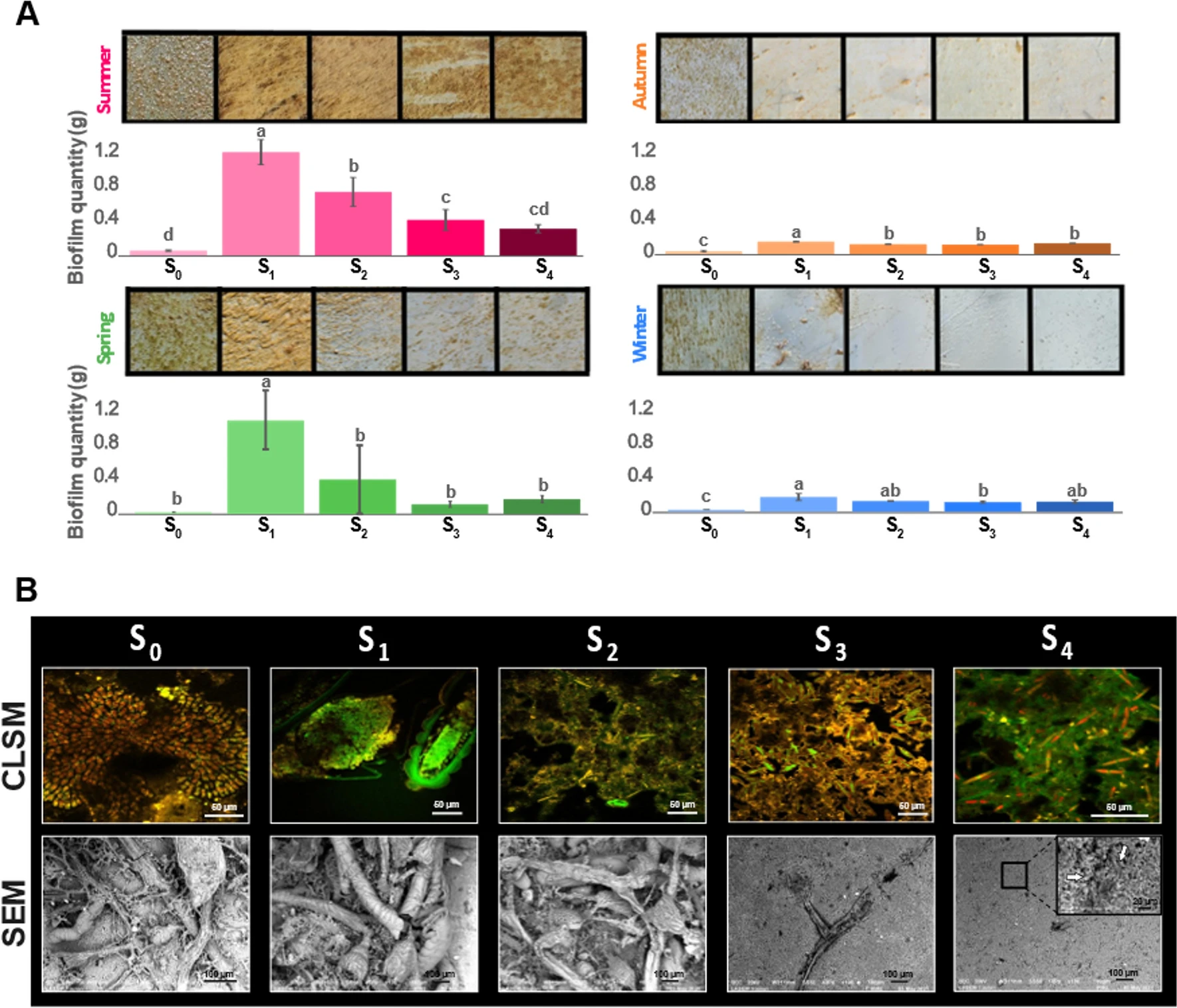
Portas et al. han estudiat la formació de biofilms sobre panells de PVC de 5 x 5 cm submergits durant 20 dies a la Badia de Toló sota un gradient de tensió tallant
Els biofilms marins com a comunitats multitròfiques
Aquest estudi fou dissenyat i concebut per Aurélie Portas (France Energies Marines, MAPIEM), Gérald Culioli (MAPIEM, IMBE), Nolwenn Quillien (FEM) i Jean-François Briand (MAPIEM). Portas realitzà els experiments, recollí les dades, va fer l’anàlisi estatística i la visualització, redactà el manuscrit original, i edità i revisà l’article. Culioli, Quillien, Briand i Annick Ortalo-Magné (MAPIEM) editaren i revisaren l’article. Nathan Carriot (MAPIEM) va fer l’anàlisi metabolòmica. Houssam Hajjoul (CNRS-IRD-MIO) i Raphaëlle Barry-Martinet (MAPIEM) van fer l’anàlisi de microscòpia confocal. Bruno Dormoy (LASEM) va fer l’anàlisi de microscòpia electrònica.
Els autors agraeixen S. Greff (IMBE) l’accés a la plataforma metabolòmica regional MALLANAR, amb seu a la Universitat d’Ais-Marselha. La recerca s’ha finançat amb el programa ANR-10-IED-0006–32 en el marc del projecte ABIOP +.
L’article fou tramès a la revista Environmental Microbiome el 9 de juliol, que l’acceptà el 21 de novembre, i el publicà el 18 de desembre.
Els biofilms o biopel·lícules són comunitats multròfiques amb alta diversitat filogenètica, que inclou microorganismes procariòtics i eucariòtics, tots ells inclosos dins d’una matriu de substàncies polimèriques extracel·lulars produïdes per una part dels microorganismes que els integren.
En ambients marins, la formació de biofilms ofereix protecció a organismes que difícilment proliferarien tant en condicions planctòniques. En el desenvolupament de biofilms influeixen factors com la disponibilitat de nutrients, la salinitat, els depredadors, la temperatura, la profunditat, el tipus de substrat o superfície, i les forces hidrodinàmiques generades per onades, corrents i marees.
En termes generals, si la hidrodinàmica genera una baixa o nul·la tensió tallant sobre la superfície submergida, el biofilm que en resulti serà menys dens, més porós però també més gruixut. En aquesta mena de biofilm es fa més senzill el transport de nutrient i l’activitat biològica.
En canvi, si la tensió de tall és molt elevada, els biofilms tendeixen a ser més compactes i densos i amb una morfologia més homogènia. L’estrès per la tensió de tall estimula la producció d’exopolímers. Però si les forces de tensió tallant superen els 3 Pa, els biofilms poden arribar a deformar-se i a desprendre’s de la superfície.
En ecosistemes marins els moviments caòtics de l’aigua tenen dos components: a) forces normals o perpendiculars a la superfície, que es poden representar amb camps de pressió; i b) forces tangencials, que són les que indueixen l’estrès per tensió tallant.
Entendre aquestes forces hidrodinàmiques és essencial per protegir superfícies artificials (sistemes d’energia renovable marina, sensors subaquàtics, cascs de vaixells, microplàstics) de la bioincrustració.
Experiments realitzats entre l’agost del 2020 i el maig del 2021
L’experiment de camp es dugué a terme a la Badia de Tolon entre l’agost del 2020 i el maig del 2021. La superfícies submergides eren panells de clorur de polivinil (PVC) tacats de sorra. Quedaven a 1 metre per sota de la superfície.
Per a les immersions dinàmiques es feien servir 18 panells de 25 x 5 cm, situats a diferent distància del centre de l’eix de rotació d’un tambor. La rotació del tambor anava a 2,7 m·s-1, de manera que les velocitats de tall anaven de 0 a 2,7 m·s-1, amb un gradient tal que S1 = 0,31 m·s-1, S2 = 0,98 m·s-1, S3 = 1,58 m·s-1 i S4 = 2,26 m·s-1.
Per a les immersions estàtiques (S0) 4 panells de 21 x 29,7 cm i 4 panells de 5 x 5 cm eren immersos a una distància suficient del rotor.
Les estructures tridimensionals dels biofilms eren observades amb un microscopi d’escaneig làser confocal (CSLM). Com a fluoròfors empraven SYTO 61 (que marca l’ADN), concanavalina A (que marca polisacàrids) i FITC (que marca proteïnes).
També s’obtingueren imatges de biofilms in situ amb l’ús d’un microscopi electrònic d’escaneig de baix buit (LV-SEM).
Per a cadascuna de les cinc condicions experimentals (S0 – S4) es recolliren biofilms de tres panells, és a dir per triplicat, amb l’ús d’un escalpel estèril, que eren col·locades en tubs de 50 mL, mantinguts a -20 °C fins a l’extracció d’ADN. Hom tenia cura de rentar prèviament amb aigua marina estèril cada panell per eliminar organismes que no formessin part de la bioincrustració.
Sobre aquestes mostres de biofilm es feia l’extracció d’ADN amb el kit Qiagen DNA Power Biofilm. L’extracte d’ADN era amplificat per a tres fragments gènics: a) 16S rRNA; b) 18S rRNA; c) gen mitocondrial de la citocrom oxidasa I (COI). Els 180 amplicons resultants eren seqüenciats amb la plataforma Illumina MiSeq, i les seqüències publicades a NCBI BioProject sota l’entrada PRJNA1052431.
L’anàlisi bioinformàtica feia servir els programes Cutadapt 3.1, DADA2 1.20 i FROGS 4.0.1. L’assignació taxonòmica es feia amb SILVA 138.2 (16S rRNA), PR2 4.14.0 (18S rRNA) i BOLD 2023 (COI).
L’anàlisi metabolòmica es feia per cromatografia líquida acoblada amb espectrometria de masses (LC-MS).
Alíquotes de biofilms eren sotmeses a un procediment d’extracció de biopolímers. Això passava per una sèrie de centrifugacions i precipitacions successives. La fracció col·loidal (CF) i la fracció lleugerament adherida (LBF) eren liofilitzades, pesades i sotmeses a una anàlisi de composició bioquímica per mètodes colorimètrics. La biomassa total s’estimava com la suma de CF, LBF i fracció residual (RF).
Les dades de seqüència genètica (metabarcoding) i de LC-MS (metabolòmica) eren combinades en una anàlisi multiòmica. Foren seleccionats 25 metabòlits, 44 seqüències de 16S rRNA, 24 seqüències de 18S rRNA i 13 seqüències de COI com a conjunt òptim de variables per a la discriminació de mostres segons si eren estàtiques o dinàmiques.
Les condicions experimentals
La temperatura dels experiments anà de 11,4 °C a l’hivern fins a 21,6 °C a l’estiu. La salinitat era de 38 i el pH de 8. La concentració d’oxigen dissolt anava del 86% (hivern) al 96% (estiu i primavera) de saturació. La concentració de nitrogen (nitrats i nitrits) era més alta a la primavera. La concentració de fosfat ho era més a l’hivern. El valor de carboni orgànic dissolt (DOC) anava de 133,95 ± 77,40 a la tardor fins a 174,69 ± 116,26 a la primavera.
Biomassa i aspecte dels biofilms segons l’estrès per tensió tallant
La comparació entre panells de diferents estacions mostra com la biomassa en l’estiu és significativament superior a la de la tardor i l’hivern.
La comparació entre panells sotmesos a diferents condicions hidrodinàmiques (S0 – S4) indica que la biomassa hi respon d’una manera dual. El valor de biomassa en panells S1 és significativament superior als panells S0 (estàtics). Però si hom compara la sèrie S1-S4, la biomassa és superior a S1 i disminueix gradualment com més gran és l’estrès per tensió tallant, de tal manera que la biomassa S4 s’acosta a la biomassa S0.
Les electromicrografies mostren un canvi entre les bioincrustracions sota baix estrès hidrodinàmic (S0-S2) respecte les d’alt estrès (S3-S4). Els biofilms de panells S3-S4 tenen un aspecte més pla, i consisteixen principalment en bacteris i diatomees inclosos en una matriu de polímer extracel·lular.
Les microfotografies amb fluorocroms indiquen que en els panells S0 trobem un biofilm dominat per estructures esfèriques tenyides de vermell per Syto61 (que correspondrien a cèl·lules) i per estructures tenyides de verd per FITC (corresponents a parets cel·lulars i components proteics). En els biofilms de panells S3-S4 s’hi distingeix una capa fina rica en diatomees (amb una clorofil·la de cloroplasts tenyida de vermell i un rafe tenyit de verd) inclosa en una matriu de biopolímer integrat per polisacàrids (tenyits de taronja) i proteïnes (tenyides de verd).
La composició de les comunitats microbianes
En les diferents estacions de l’any trobem als panells una molt lleugera alteració en la composició procariòtica dels biofilms, estimada d’acord amb el 16S rRNA. Més rellevant són els canvis observats segons les condicions hidrodinàmiques en els panells d’hivern i de primavera.
L’índex de Shannon és un indicador matemàtic de diversitat. Pel que fa als priocariotes varia entre els panells S0 i S4, excepte en l’estiu quan no ho fa.
Pel que fa a la diversitat d’eucariotes, valorada per 18S rRNA, l’índex Chao 1 mostra que en els panells de tardor S1 indueix un augment de diversitat respecte dels panells S0 de la mateixa estació.
Pel que fa a la diversitat mesurada per COI, aquesta disminueix en S1-S2 respecte de S0, però torna a augmentar en S4 en l’estiu i l’hivern. A la tardor, la diversitat COI és superior en S1-S4 respecte a S0. A la primavera, la diversitat COI és inferior en S1-S4 respecte a S0.
En resum, la introducció d’un estrès per tensió tallant té un impacte sobre la composició de la comunitat microbiana superior al canvi que pugui induir un augment de la intensitat d’aquest estrès. Altres factors que influeixen en la diversitat mesurada per rRNA són la temperatura, la concentració de nitrit, la concentració de nitrat i el pH. Pel que fa a la diversitat COI hi influeix la salinitat, la concentració de silicats, la concentració de nitrits i la concentració de nitrats.
L’estrès per tensió tallant afecta més les comunitats procariòtiques que no pas variacions en paràmetres ambientals. En comunitats eucariòtiques, l’efecte de l’estrès per tensió tallant no és tan marcat.
En aquests experiments les comunitats de procariotes més abundants són les Rhodobacteraceae, les Flavobacteriaceae, les Pseudomonadaceae i les Vibrionaceae. A l’estiu i la tardor hi dominen Flavobacteriaceae i Saprospiraceae, mentre que a l’hivern i a la primavera ho fan les Rhodobacteraceae.
El gènere procariòtic més abundant en condicions estàtiques (S0) és Winograskella, i ho és en totes quatre estacions. En condicions dinàmiques (S1-S4), els gèneres més abundants són Tenacibaculum i Aquibacter, amb un caiguda en la tardor. En la tardor i hivern, en els panells S1-S4 hi ha un augment de l’abundància de Sulfurovum i Mycoplama.
La combinació de dades 18S rRNA i COI ofereix un retrat de la comunitat eucariòtica. Aquesta comunitat eucariòtica varia amb les estacions de l’any. Així, les Dinophyceae són presents únicament en els panells dinàmics de la primavera. Entre els hidrozous, Bougainvillia és dominant a la tardor i l’hivern en panells dinàmics, succeïda en la primavera per Obelia en la primavera i hivern, excepte en panells S4 quan hi fan presència Hydractinia i Coryne.
En panells estàtics trobem indicis de presència d’artròpodes (copèpodes i malacostracis), nematodes (Chromadorina), rodòfits (Ceranium) i mol·luscs (Eubranchus).
En panells dinàmics de primavera trobem un predomini de diatomees i de Dothiomycetes (Phaosphaeria i Toxicocladosporium).
Organismes específicament associats amb les condicions d’estrès per tensió de tall
Hi ha una sèrie de seqüències gèniques comunes a panells estàtics i dinàmics. D’altres són específics dels panells dinàmics (com el rodobacteriaci Salinihabitans; els ciliats Euplotes, Loxophyllum i Disteria; els briozous Watersipora i Scuparia).
Canvis metabolòmics
L’anàlisi de components principals mostra que les condicions hidrodinàmiques altera el metaboloma dels biofilms. El dimetilsulfonipropionat (DMSP) és un quimiomarcador de panells dinàmics. El liso-DGTA i àcids grassos simples són més abundants en panells estàtics.
Canvis en biopolímers extracel·lulars
Els panells dinàmics de l’hivern i de la tardor són els presenten menys quantitat de biofilm. En aquestes condicions els biopolímers són molt variables, i cauen en els panells S2-S4 respecte de S1.
Polisacàrids i àcids urònics s’associen amb panells estàtics en totes les estacions de l’any. Les proteïnes s’associen amb panells dinàmics de baix estrès.
Anàlisi multiòmica
L’anàlisi combinada de genoma i metaboloma mostra dos clústers:
– un associat amb panells estàtics integrat per procariotes com Ochrovirga, Erythrobacter i Winogradskyella i per metabòlits com feofòrbid a, nitenina i estigmatel·lina Y.
– un associat amb panells dinàmics.
Hidrodinàmica i bioincrustració
Portas et al. mostren com la intensitat de la tensió tallant no es relaciona linealment ni amb la biomassa dels biofilms ni amb la producció de biopolímers extracel·lulars. D’entrada, una major temperatura, en promoure la productivitat primària de l’aigua marina, estimula un major creixement de biofilms en l’estiu. Un cert nivell de tensió de tall maximitza alhora la biomassa del biofilm, però ultrapassat aquest llindar l’efecte és negatiu. És possible que l’efecte beneficiós d’una tensió de tall moderada es degui a un augment de la disponibilitat de nutrients i d’oxigen, però també pot constituir un obstacle a la depredació exercida per espècies vàgils.
En panells S2-S4, l’estrès de tensió de tall comportaria una caiguda de la biomassa dels biofilms en limitar l’adhesió a la superfície. El biofilm és més prim, però alhora hi ha un augment de la producció d’expolisacàrids. Aquest augment s’explicaria com una resposta adaptativa del biofilm, que guanya així en cohesió. De tota manera, en la condició S4 hi ha una disminució notable de la diversitat procariòtica del biofilm.
En condicions d’alt estrès de tensió tallant, els microorganismes dels biofilms han d’ésser capaços d’ancorar-se a la superfície, bé a través d’estructures com pili o curli, o de la secreció de biopolímers. Espècies colonials d’ascidis, hidrozous i briozous tenen capacitat d’incrustració en aquestes condicions dinàmiques.
En condicions estàtiques hi ha una major abundància d’algues brunes (Leathesia, Pylaiella) i vermelles (Ceramium). També hi ha major presència del nematode Chromadorina i del gastròpode Eubranchus, i en general de la fauna vàgil representada per copèpodes i amfípodes.
El cas del dinoflagel·lat Biecheleria il·lustra sobre el rol que els biofilms marins poden jugar en ecosistemes litorals. Biecheleria no es troba pas en l’ambient planctònic de la Badia de Tolon, però en canvi és abundant en els biofilms d’aquest estudi. Els biofilms ofereixen hàbitat a organismes que són rars en el plàncton, com passa amb algunes espècies del gènere Vibrio que poden causar mortaldats en explotacions aqüícoles (V. owensii, V. crassostreae o V. campbelli). Quelcom semblant passa amb algunes espècies invasores com Watersipora subtorquata i Burgula neritinia, detectades en panells de tardor i d’hivern d’aquest estudi.
Lligams:
– Shear stress controls prokaryotic and eukaryotic biofilm communities together with EPS and metabolomic expression in a semi-controlled coastal environment in the NW Mediterranean Sea. Aurélie Portas, Nathan Carriot, Raphaëlle Barry-Martinet, Annick Ortalo-Magné, Houssam Hajjoul, Bruno Dormoy, Gérald Culioli, Nolwenn Quillien, Jean-François. Briand. Environmental Microbiome 19: 109 (2024).
– Study of the biofilm on renewable marine structures. Aurélie Portas (2023). http://www.theses.fr/s227947
