Sota el nom de baldriga, en català, o de shearwater, en anglès, es coneix un ample ventall d’ocells marins de la família dels procel·làrids. Tenen en comú un comportament pelàgic, i de fet tan sols abandonen la mar per tal de criar en el litoral. Disposen d’un gran sentit de l’orientació, que els ajuda a retornar a la zona de cria però també en la recerca de recursos alimentaris. Com s’ho fan per distingir superfícies marines immenses, sempre canviants i aparentment desprovistes de senyals diferenciadores? Experiments basats en la inducció d’anòsmia en baldrigues indiquen que el sentit de l’olfacte és essencial per tal de tornar a l’àrea habitual des d’un àrea allunyada. Però des de l’Oxford Navigation Group, Tim Guilford i Ollie Padget són conscients de les dificultats d’interpretar aquests experiments de pertorbació sensorial (olfactiva o magneto-receptora), en tant que les capacitats de navegació poden veure’s confoses amb altres factors. Així Padget i Guilford pensaren en un disseny més robust. Decidiren aplicar-lo a la baldriga cendrosa mediterrània (Calonectris diomedea), i comptaren amb el suport d’Anna Gagliardo, de la Universitat de Pisa, i de Gaia Dell’Ariccia i Jacob González-Solís, de l’Institut de Recerca de la Biodiversitat, de la Universitat de Barcelona, per fer el treball de camp. Els resultats d’aquest estudi, que es publica a Scientific Reports, indiquen que l’anòsmia afecta l’orientació de retorn però no pas les capacitats depredadores.

L’espècie Calonectris diomedea fou descrita en el 1769 per Johannes Antonius Scopoli. Per això se la denomina “baldriga d’Scopoli”, si bé la seva relació taxonòmica amb altres baldrigues és encara matèria de discussió. Entre els noms comuns que rep en català hi ha, a més de “baldriga cendrosa mediterrània” el de “baldritja gran” o el de “virot gros”. En la imatge, captada el 18 de maig del 2012 al Parc Nacional de Port-Cros e Pòrcaròlas, a Provença, hi veiem un exemplar
Un experiment a Cala Morell, l’estiu del 2016
Nascuda a Pisa el 3 de maig del 1957, Anna Gagliardo es va doctorar en etologia a la Universitat d’aquesta ciutat en el 1989. Des del 2004 és investigadora al Departament de Biologia de la Universitat de Pisa, on participa en la docència de “Bases neutrals del comportament”. El seu interès principal és conèixer la natura dels estímuls sensorials en els quals es fonamenta el mapa de navegació dels ocells. Gagliardo et al. (2013) feren un important experiment de desplaçament en la baldriga atlàntica de Cory (Calonectris borealis). En aquest experiment, baldrigues que crien a les Açores foren capturades i se les transportà a una distància de 800 km en l’Oceà Atlàntic. A algunes d’elles se les havia induït una anòsmia, exposant-les a un rentat de mucosa olfactòria amb sulfat de zinc, i a d’altres se les pertorbà el sentit magnètic col·locant-les magnets mòbils al cap. Les baldrigues del grup control i del grup dels magnets (amb una excepció) retornaren a les Açores, però de les vuit baldrigues anòsmiques tan sols tornaren a casa dues.
Guilford i Gagliardo participaren en un altre estudi, aquest realitzat sobre poblacions de l’arxipèlag toscà de la baldriga d’Scopoli (Pollonara et al., 2015), però amb els mateixos procediments experimentals. Els ocells eren alliberats en aquest cas a mar obert. Les baldrigues anòsmiques tornaren a casa però ho feren més tard que el grup control i el grup dels magnets.
Els experiments de desplaçament permeten reconèixer els elements que participen en la navegació de retorn. En canvi, per a la navegació habitual espontània dels ocells es fan altres tipus d’estudis, i en aquests no s’ha estudiat com hi afecta l’anòsmia.
Ollie Padget i Tim Guilford, doncs, dissenyaren un experiment per saber si l’anòsmia induïda per sulfat de zinc o la disrupció magnètica afecten els desplaçaments naturals de les baldrigues. Per seguir els moviments dels ocells empraren petits registradors de geolocalització.

Cala Morell, al nord del terme municipal de Ciutadella. A Menorca és on trobem les colònies més importants de baldritja gran de la Mediterrània, si bé és a la Macaronèsia on hi ha la població més nombrosa. A Menorca és present tot l’any, si bé és a partir de març i fins a l’estiu, abans del capvespre, quan sovinteja més a la costa (cap de Favàritx, Cavalleria, Punta Nati).
Padget, Dell’Ariccia, Gagliardo, González-Solís i Guilford monitoritzaren la colònia de baldritja gran (o d’Scopoli) entre el 25 de juny i el 21 de juliol del 2016. Seguien el darrer terç del període d’incubació dels ous, i identificaren les parelles nidificants que podien prendre per a l’estudi. Ho feien en el marc d’un estudi més general de la colònia, on es capturen els nous ocells, s’anellen i es prenen dades morfomètriques. Els ocells són capturats manualment o amb un llaç.
Un total de deu ocells foren fets anòsmics amb un rentat de la mucosa olfactiva a través dels narius amb 4 ml d’una solució aquosa al 4% de ZnSO4·7H2O. Aquest tractament produeix la necrosi de les neurones de la mucosa olfactiva, anul·lant el sentit de l’olfacte fins que noves cèl·lules de la mucosa es diferencien i connecten amb el bulb olfactori. En coloms, que és on s’ha estudiat aquest procés, la recuperació de l’olfacte triga força setmanes.
Un altre grup de deu ocells fou sotmès a una disrupció magnètica. Se’ls col·locava entre els ulls un magnet cilíndric (4×5 mm) de neodimi (Nd2Fe14B).
Com a grup control s’emparen dotze ocells. A aquest grup control, com al grup anòsmic, se’ls col·locava, per comptes del magnet de neodimi, una boleta de vidre de dimensions similars.
Als 32 ocells de l’estudi, se’ls havia col·locat, just abans del tractament corresponent, dispositius de GPS, un per cada ocell, situat en les plomes de l’esquena. Cada GPS pesa 18 g, que es correspon a un 3% de la massa corporal d’una baldritja gran (580 g); i té unes dimensions de 9x3x1,5 cm.
Totes aquestes manipulacions havien estat aprovades pel Comitè Ètic de la Universitat d’Oxford i seguien la normativa de recuperació del virot del Decret 65/2004 del Govern de les Illes Balears. En quatre dels ocells participants es comprovà que el tractament no impactés negativament en el comportament d’incubació. El temps de manipulació de cada animal, comptant la inicial i la de recuperació, era normalment inferior a 15 minuts.
Seguiment de GPS cada 5 minuts
Cada GPS registrava la seva posició cada 5 minuts. Quan eren recuperats pels investigadors, s’analitzava el recorregut realitzat. Normalment, cada registre contenia múltiples viatges. Mitjançant l’algoritme de segmentació de Douglas-Peucker les línies crebades obtingudes eren traduïdes a viatges individuals. En cada viatge individual també identificaven el moviment corresponent al retorn al niu. Es considerava una distància de 40 km de l’arxipèlag balear per distingir entre la zona costanera i la pelàgica. Aplicaren el criteri del crepuscle nàutic (quan el Sol és a més de 6º per sota de l’horitzó) per definir la part nocturna dels desplaçaments.
Les dades de moviment obtingudes per Padget et al. han estat dipositades al Movebank.
Com que tant en la captura inicial (de 29 de juny a 10 de juliol) com en la recuperació del GPS (de 6 de juliol a 20 de juliol), els animals eren pesats, s’obtenia així una dada del canvi de massa corporal. Com que hi ha una pèrdua de massa corporal associada a la incubació dels ous, Padget et al. feren un ajustament basat en dades d’uns altres vuit ocells.
Els resultats
Dels 32 dispositius de GPS, va haver-hi un, col·locat en un ocell anòsmic, que es va quedar sense bateria durant l’experiment. En conseqüència, els grups quedaren constituïts per 12 ocells control (C), 10 ocells manipulats magnèticament (M) i 9 ocells anòsmics (A). Aquests 31 ocells, en el decurs de 15 dies d’experiment, feren un total de 59 viatges (16 C; 23 M i 20 A). Els 32 ocells manipulats, després d’haver estat alliberats del GPS, tornaren a incubar els ous, i feren nous viatges, dels quals tornaren tots abans de 21 de juliol excepte un d’ells, que havia estat en el grup M.
Dels 32 nius d’on s’obtingueren els 32 ocells manipulats, eclosionaren els polls en 18 casos. En 14 no ho feren (4 del grup C, 6 del grup M i 4 del grup A), principalment perquè l’ou fou depredat per la rata bruna (Rattus norvegicus). En les baldritges és habitual que sempre hi hagi un progenitor en els nius, però això no sempre es compleix, i en els nius de l’estudi, va haver-hi 6 que quedaren desprotegits durant 1-3 dies.
El sistema de subjecció de magnets (boles de vidre en els grups C i A) no era gaire efectiu. De fet, en el moment de la recuperació només duien aquests dispositius 11 dels 31 ocells.
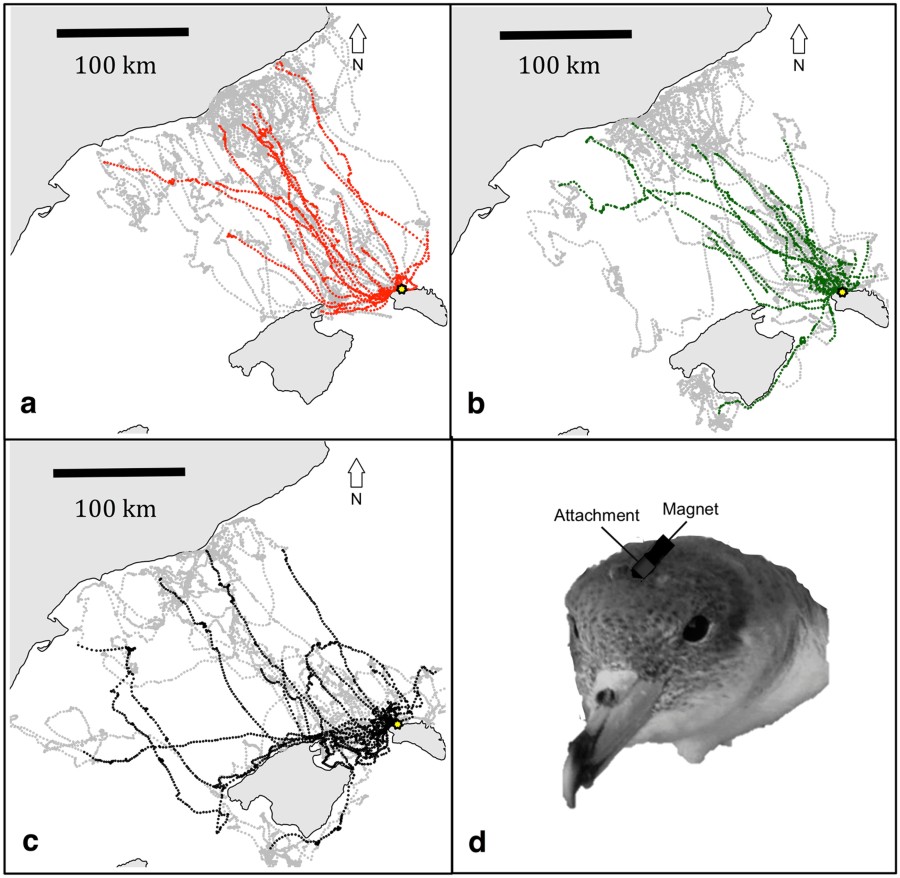
Els 59 viatges realitzats pels 31 ocells de l’estudi, dividits entre el grup control (a), magnètic (b) i anòsmic (c). En cada mapa s’assenyala en gris el viatge d’anada i en color (vermell en el grup C; verd en el grup M; negre en el grup A) el viatge de tornada.
Els tres grups no es diferenciaven ni en el temps que trigaven fins a fer el primer viatge des de la Cala Morell. Tampoc no hi havia en el nombre total de viatges realitzats entre els grups.
Els 59 viatges de l’estudi eren típicament llargs, amb una durada de 3 a 10 dies. Cada viatge té una fase d’allunyament del litoral menorquí i una fase d’acostament, i entre mig hi ha un llarg període que transcorre bé davant del litoral continental (20; entre el Delta de l’Ebre i les Muntanyes de Begur), davant del litoral balear (11) o en zones pelàgiques de la Mar Balear (27). En aquestes àrees, les baldrigues s’alimenten de petits peixos i cefalòpodes, que capturen en la superfície o en petites capbussades.
No hi havia diferències entre els tres grups quant a la preferència regional, la durada del viatge (100 hores de mitjana) o la distància coberta (700-1100 km).
Dels 59 viatges es poden inferir tres estats:
– períodes en els quals l’ocell reposa en l’aigua, amb velocitats mitjanes de 0,3 m/s.
– períodes de vol dirigit, amb velocitats mitjanes de 6,7 m/s.
– períodes de vol de predació, amb velocitats mitjanes de 1,8 m/s.
Tampoc no hi havia diferències entre els grups quant a la proporció d’aquests tres estats de velocitats diferenciades, ni tampoc quant a la proporció dels estats entre el dia i la nit. En tots tres grups, els períodes de repòs es concentren en la nit, mentre que els vols dirigits es realitzen en hores crepusculars (tant d’albada com d’ocàs).
Pel que fa al canvi de massa corporal, en general els ocells tendien a guanyar massa, però no es registraven diferències entre els tres grups tampoc en aquest punt.
En els 59 viatges analitzats, l’anada segueix una orientació similar entre els tres grups. Ara bé, el grup anòsmic triga una mica més en allunyar-se de la costa.
Una de les capacitats de navegació es manifesta en el temps de retorn d’un viatge. Els ocells calculen l’inici del viatge de retorn per poder arribar al niu a la nit. Com més distant s’hi troben inicialment, abans hauran de començar el viatge. Aquesta capacitat es manté similar en els tres grups.
Una altra capacitat es manifesta en els ocells que es troben en àrees pelàgiques i que emprenen el camí de retorn. En aquest cas sí hi ha diferències. Mentre el grup C i M mostren una bona orientació, no és pas tan bona la dels ocells anòsmics. Els tres grups segueixen línies més o menys rectes, però els anòsmics ho fan en una orientació generalment equivocada.
Si l’orientació pelàgica minva en els ocells anòsmics, no s’esdevé el mateix en l’orientació litoral. Els tres grups manifesten orientacions similars de tornada quan aquesta es produeix des de la zona litoral balear.
L’olfacte en l’orientació pelàgica
Padget et al. mostren com ni la pertorbació de l’olfacte ni de la magnetorecepció alteren la capacitat de les baldrigues per iniciar viatges, depredar-hi de manera prou exitosa com per guanyar pes i retornar-hi per continuar la incubació. És clar que les conclusions de la magnetorecepció es veuen afectades negativament pel fet que en la majoria de casos, els magnets s’havien ja desprès dels ocells en el moment de la recaptura.
L’anòsmia induïda, doncs, no sembla afectar ni la capacitat de predació ni els patrons de comportament en la zona pelàgica. Això és rellevant en la interpretació dels estudis de desplaçament on es temia que l’anòsmia afectés altres funcions no relacionades amb la navegació.
L’anòsmia tampoc no sembla alterar processos cognitius subjacents a la navegació com ara el càlcul del temps necessari per fer el viatge de retorn. Tampoc no altera la capacitat de fer un vol directe en línia recta, probablement perquè això depèn d’una detecció de l’orientació geogràfica general (brúixola).
Ara bé, l’anòsmia sí afecta a l’orientació en la zona pelàgica per fer el camí de retorn. Els animals anòsmics trien una ruta generalment equivocada, i no la corregeixen fins que no arriben a la zona litoral (<40 km).
Per Padget et al., la informació olfactiva és rellevant sobretot en la navegació de retorn cap a la colònia una vegada finalitzada l’etapa depredadora d’un viatge. Així, el “mapa olfactiu” no és indispensable per al moviment de les baldrigues però sí és consultat en arribar el moment de tornar a la colònia. Fins ara, això s’havia vist en estudis de desplaçament forçat, però Padget et al. ho mostren en l’autodesplaçament.
El “mapa olfactiu” d’alguna manera és essencial per aconseguir una orientació correcta en la zona pelàgica en la fase de tornada. Una vegada s’arriba a la fase litoral, el mapa olfactiu cedeix protagonisme al “mapa visual”.
És interessant que l’olfacte no sembli crucial perquè les baldrigues trobin aliment en el seu viatge. No obstant, Dell’Ariccia et al. (2014) assenyalaren que les baldrigues localitzen riquesa de preses a través d’estímuls olfactius (se senten atretes per l’olor de sulfur de dimetil). Potser els ocells anòsmics recolzen en la seva memòria o en senyals visuals (topografia, congregació d’altres ocells marins, etc.).
En qualsevol cas, Padget et al. consideren que el més rellevant d’aquest estudi és mostra l’efecte específic en la navegació de l’anòsmia induïda per sulfat de zinc en un ocell marí.
Lligams:
– Anosmia impairs homing orientation but not foraging behaviour in free-ranging shearwaters. O. Padget, G. Dell’Ariccia, A. Gagliardo, J. González-Solís, T. Guilford. Scientific Reports 7, 9668 (2017).
