En la determinació del rang geogràfic d’una espècie, hom assum que hi són determinants factors ecològics, i que el rang geogràfic s’acaba allà on l’espècie topa amb el marge ecològic que li és propi. En les zones marginals, la duresa de les condicions posa a prova la plasticitat de l’espècie, i és justament en aquests indrets on s’esperaria trobar una major variabilitat fenotípica. En el cas de les plantes, aquesta variabilitat és fa més evident en els trets vegetatius (arrels, tija, fulles) que no en els trets reproductius (flors, fruits), ja que la pressió selectiva contra la variabilitat és més forta en aquests darrers. Carmelo Macrì, de la Universitat de Gènova, i Gabriele Casazza, de la Universitat d’Ais, juntament amb altres investigadors del projecte ADCLICH utilitzen Lilium pomponium, un llir endèmic dels Alps Lígurs i Marítims com a model de variació morfològica i genètica a través d’un gradient ambiental. En un article a la revista Oecologia expliquen que hi ha una relació entre la variació de trets florals i les condicions ambientals locals, mitjançada per factors com la disponibilitat de recursos, la limitació potencial del pol·len i la mida de la població. De tota manera, Macrì et al. troben que tant les poblacions centrals com les perifèriques tenen uns valors similars de producció de llavors, encara que hi arriben amb diferents estratègies de cicle vital. No semblaria, doncs, en el cas d’aquesta espècie, que les diferències en l’ambient de pol·linització siguin el principal determinant del límit de distribució.
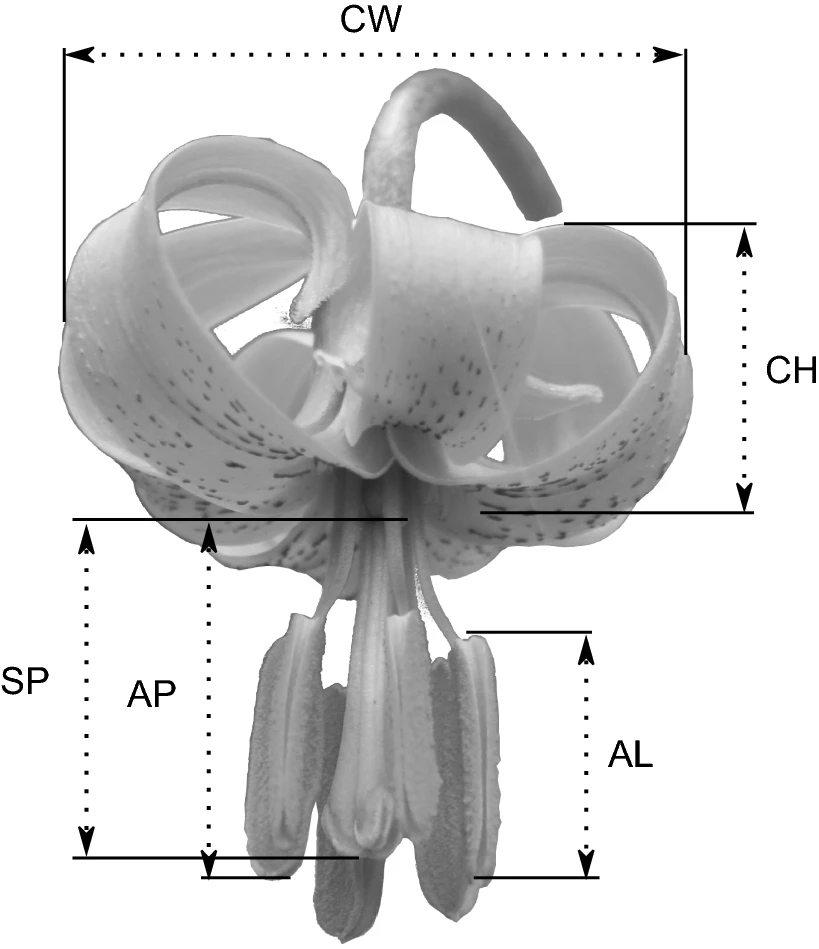
En termes generals, els trets florals són més constants que trets vegetatius de les plantes. És per això que en aquest estudi sobre “Lilium pomponium”, Macrì et al. quantifiquen paràmetres florals, com l’amplada de la corol·la (CW), l’alçada de la corol·la (CH), la posició de l’estigma (SP), la posició de l’estam (AP) i la longitud de l’estam (AL). Es tracta de reconèixer variacions en aquests trets d’acord amb la situació de la localitat en el rang geogràfic de l’espècie
La hipòtesi del centre-perifèria
La idea d’aquest estudi fou formulada per Carmelo Macrì, del Dipartimento di Scienze della Terra dell’Ambiente e della Vita (DISTAV), de la Universitat de Gènova, i per Gabriele Casazza, de l’Institut Méditerranéen de Biodiversité et d’Ecologie marine et continentale (IMBE), amb seu a Ais de Provença. En el desenvolupament metodològic i el treball de camp participaren Macrì, Davide Dagnino (DISTAV de Gènova), Luigi Minuto (DISTAV de Gènova) i Casazza. Les anàlisis estatístiques foren conduïdes per Macrì i Casazza. En la redacció de l’article participaren Macrì, Dagnino, Minuto, Frédéric Médail (de l’IMBE), John D. Thompson (del Centre d’Ecologie Fonctionelle et Evolutive, de Montpelhièr) i Casazza. Els autors agraeixen la col·laboració en el treball de laboratori i de camp de Sara Nicolini, Chiara Calise, Alessandro Rivata i Aleandra Di Caro.
El treball fou finançat amb el projecte Horitzó 2020 concedit a ADCLICH. Minuto fou l’autor corresponsal davant de la revista Oecologia, que rebé el treball l’11 de desembre del 2019. Després d’un llarg procés de revisió, l’article fou acceptat el 12 de novembre del 2020, i publicat el 2 de desembre.
El punt de partida és la hipòtesi centre-perifèria, segons la qual les poblacions perifèriques i aïllades geogràficament d’una espècie haurien de divergir de les poblacions centrals, de tal manera que les poblacions perifèriques serien més petites, menys abundants i més aïllades entre elles. Les poblacions perifèriques s’enfronten a una qualitat d’hàbitat inferior i, en el cas de les plantes, hom pensa que el factor cabdal consistiria en una reducció de la qualitat i quantitat de la pol·linització.
El trasllat del concepte de centre-perifèria es fa difícil en ecosistemes alpins mediterranis, que mostren una gran heterogeneïtat, vinculada a la complexitat topogràfica del relleu alpí.
L’espècie d’estudi de Macrì et al. és Lilium pomponium L. S’hi fixen en els caràcters florals, ja que en principi són els més conservats en diferents condicions ambientals. L. pomponium és una planta perenne geofítica, és a dir que rebrota cada primavera a partir d’un bulb subterrani. És auto-incompatible, és a dir que no pot autofecundar-se. És una planta endèmica dels Alps Marítims i Lígurs, que creix damunt d’afloraments calcaris a altituds entre 100 i 2000 m. Dins d’aquest rang geogràfic es trobada des de zones de clima mediterrània (d’estiu eixut) a zones de clima continental (d’estiu fresc).
Macrì et al. volien comprovar si les poblacions geogràficament perifèriques de L. pomponium són també poblacions ecològicament marginals, i alhora saber si aquestes poblacions ecològicament marginals difereixen fenotípicament de les poblacions centrals en trets florals, i si aquesta diferència es tradueix en una minva de la capacitat reproductiva.
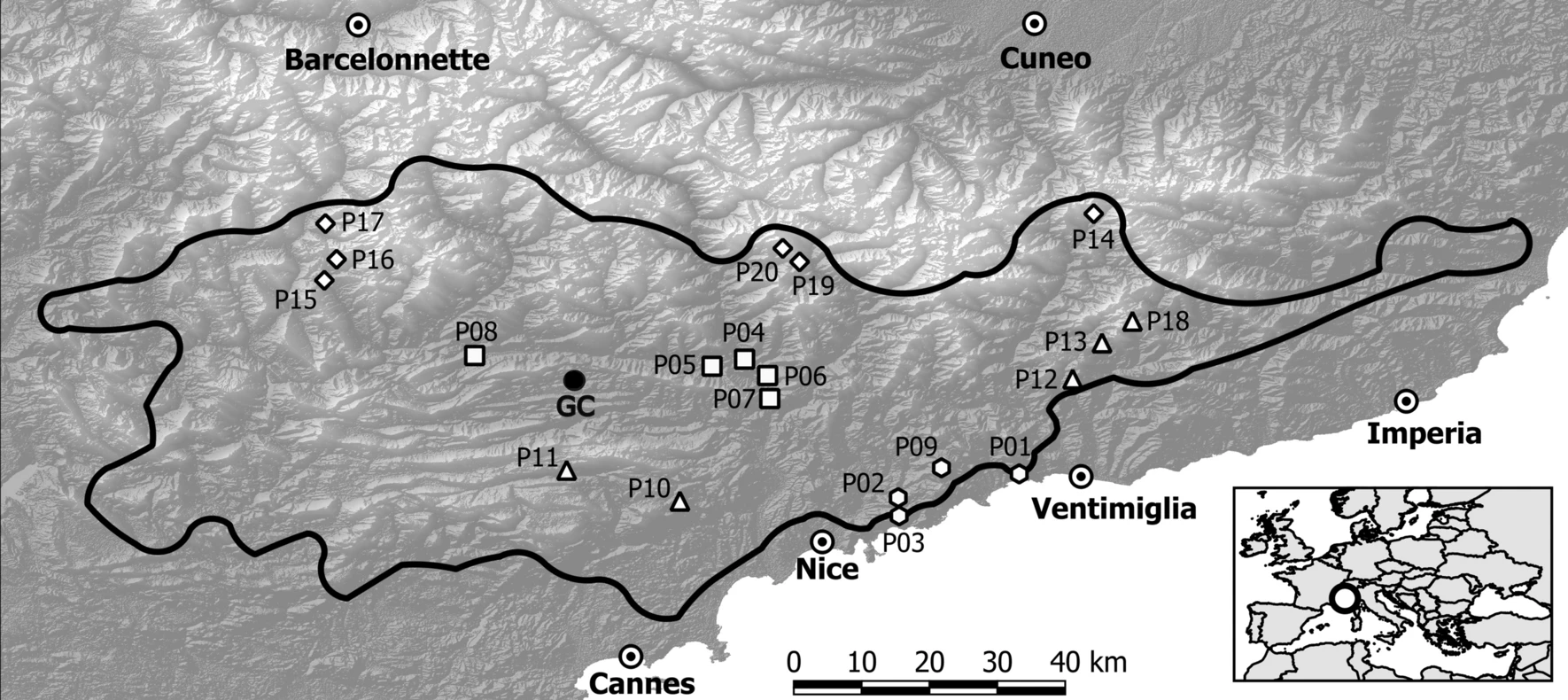
Rang de distribució de “L. pomponium” als Alps Mediterranis, on s’assenyalen les 20 poblacions emprades en l’estudi de Macrì et al. Les localitats es classifiquen en quatre grups climàtics: central (triangles), litoral marginal (hexàgons), interior marginal (rectangles) i subalpí marginal (rombs)
Vint poblacions de L. pomponium
Les flors de L. pomponium són en general hermafrodites, per bé que vora un 10% són flors exclusivament masculines. La floració va de maig a juliol, i la fructificació de final de juliol a setembre. Encara que la flor sigui hermafrodita, no pot autofecundar-se (auto-incompatibilitat), entre d’altres coses pel factor anatòmic que els estigmes femenins siguin més elevats que els estams masculins. Aquesta auto-incompatibilitat explica que la taxa de flors que arriben a fructificar sigui baixa. Les llavors es dispersen pel vent, gràcies a un marge en forma d’ala.
El rang de distribució de L. pomponium que s’utilitza en aquest estudi és el determinats pels exams de camp dels propis autors, i pels registrats a les bases de dades de SILENE i LiBiOss. Segons aquests rangs, aplicant-hi un filtre espacial d’1 km, hi hauria presència de l’espècie en 809 localitats.
Pel que fa a la informació climàtica, Macrì et al. empren dinou variables bioclimàtiques corresponents al període 1979-2013 disponibles en la base de dades CHELSA.
La distinció entre població central i població ecològicament marginal, Macrì et al. la basen en una anàlisi de components principals de les variables bioclimàtiques. Així obtenen una el·lipse de confiança del 70%, dins de la qual quedarien les poblacions “centrals”.
D’aquesta manera, poden comprovar la correlació existent entre el “centre geogràfic” del rang de distribució de l’espècie i el “centre ecològic”.
Entre el 2017 i el 2018, foren triades a l’atzar en les 40 localitats referides, 414 plantes (de 8 a 34 per cada població). D’aquestes plantes, resultaren mesures de 562 flors (250 en el 2017 i 312 en el 2018). Al mateix temps, computaven el nombre de flors per escap i la superfície de corol·la, com a factors determinants en l’atracció de pol·linitzadors. Les mesures de flors es feien en el camp mateix, amb un peu de rei digital, amb una precisió de 0,01 mm.
D’acord amb les mides d’estams i d’estigmes, les flors podien ser classificades en: 1) flors que s’acostaven a una situació d’herkogàmia, amb l’estigma per damunt de l’estam; 2) flors que mostren una situació d’herkogàmia inversa, amb l’estigma per sota de l’estam; 3) flors sense herkogàmia, amb un estigma i estams situat a la mateixa alçada.
En 12 de les 20 poblacions, es taparen amb una bossa 183 brots florals de 139 plantes diferents. D’aquesta manera, aquests brots florals únicament podrien donar lloc a fruits per autofertilització.
En 17 de les 20 poblacions, es practicà una pol·linització manual en 168 flors de 143 plantes diferents, mentre que es deixaven com a control de pol·linització oberta 344 flors de 279 plantes diferents. Posteriorment, es realitzà un recompte de les llavors presents en les càpsules madures resultants d’aquestes flors. D’aquesta manera, hom podia fer una estimació de la limitació pol·línica de cadascuna de les 17 poblacions.
Pol·linització i marginalitat
D’acord amb l’anàlisi de components principals, de les 20 poblacions analitzades, 5 serien reconegudes com a centrals i 15 com a marginals. Dins de les 15 poblacions marginals, es distingeix entre les litorals (amb condicions càlides i humides), les interiors (amb condicions càlides, però més seques) i les subalpines (amb condicions fredes i humides).
Les flors eren més grans en les zones de menys altituds, dins de les poblacions marginals costaneres i interiors. Així doncs, mentre que les flors de les poblacions marginals subalpines tenien una mida mitjana de 2,49 mm2, les poblacions marginals litorals arribaven a una superfície floral mitjana de 3,14 mm2. De manera corresponent, el nombre de flors per escap era inferior en les poblacions litorals i centrals (1-2) en comparació amb els nombres de les poblacions interiors i subalpines (2-3).
La majoria de flors analitzades (86%) no presentaven herkogàmia (els estams eren a l’alçada dels estigmes). Les poblacions costaneres eren les que presentaven un índex superior d’herkogàmia (25%).
Pel que fa als estudis de limitació pol·línica, els valors més elevats es trobaven en les poblacions marginals costaneres i interiors. En les poblacions centrals la limitació pol·línica és pràcticament zero, i en les poblacions marginals subalpines té un valor fins i tot lleugerament negatiu. El caràcter auto-incompatible de l’espècie queda ratificat pel fet que de les 93 flors a les que se’ls havia col·locat una bossa, tan sols 1 va arribar a produir fruit.
Centre geogràfic i centre ecològic
Hi ha una certa congruència entre la marginalitat geogràfica i la marginalitat ambiental de les poblacions de L. pomponium. Així les poblacions marginals costaneres es troben en l’extrem sud del rang geogràfic de distribució, i les poblacions marginals subalpines en l’extrem nord d’aquest rang. De tota manera, les poblacions marginals interiors es troben més a prop del centre geogràfic de distribució de l’espècie que les poblacions ecològicament centrals. Comptat i debatut, Macrì et al. no troben correlació entre les distàncies geogràfiques i les distàncies climàtiques. Així doncs, hom no pot, en el cas d’aquesta espècie, identificar centre geogràfic amb centre ecològic: els gradients geogràfics i ambientals no coincideixen en un relleu certament accidental.
Les poblacions marginals costaneres i interiors presenten unes flors més amples. En el cas de les poblacions marginals interiors, hi ha un major nombre de flors per escap. Les poblacions marginals, sotmeses a una limitació d’accés a agents pol·linitzadors, s’adaptarien d’aquesta manera: unes flors més grans i una densitat floral superior ajuda a atreure pol·linitzadors (lepidòpters).
No totes les poblacions marginals de L. pomponium patirien una limitació de pol·linitzadors. En les poblacions marginals subalpines la pressió selectiva seria una altra: la impredictibilitat de l’ambient afavoriria unes flors més petites, però més abundants, i amb un període de floració més llarg.
Quan Macrì et al. comparen les poblacions centrals i les marginals en la quantitat de llavor produïda, no hi troben diferències. Això podria deure’s al fet que en les poblacions centrals i subalpines, si bé hi ha una bona pol·linització, també hi ha una depredació de fruits per part del coleòpter Lilioceris lilii.
Estudis de camp com aquest no poden discriminar si les variacions en trets anatòmics són degudes a fenòmens de plasticitat fisiològica o d’adaptació evolutiva. Per això, ADLICH té prevista la realització d’experiments en hivernacles que aclareixen quina variació genotípica hi ha darrera de la variació fenotípica entre poblacions de L. pomponium.
Lligams:
– Effects of environmental heterogeneity on phenotypic variation of the endemic plant Lilium pomponium in the Maritime and Ligurian Alps. Carmelo Macrì, Davide Dagnino, Maria Guerrina, Frédéric Médail, Luigi Minuto, John D. Thompson & Gabriele Casazza. Oecologia (2020)
