Sota el nom d’algues blaves, algues cianofícies, de cianoprocariotes o de cianobacteris, hom entén una divisió de bacteris amb la sinapomorfia de posseir un metabolisme fotosintètic i oxigènic. Un llinatge de cianobacteris donà lloc als “cloroplasts” que trobem en les algues eucariòtiques i, a través d’elles, en les plantes. D’altres llinatges mantenen encara la categoria de simbionts (i no de mers orgànuls) d’altres organismes. Finalment, la majoria de cianobacteris viuen de manera lliure en la columna d’aigua (dolça o marina), o bé en el fons de llacs, rius o mars, o fins i tot en zones humides emergides. Entre els cianobacteris simbionts hi ha el gènere Phormidium i d’altres de semblants. En un article a la revista Biology Open, el grup de recerca de Valerio Zupo, de la Stazione Zoologica Anton Dohrn, de la Punta San Pietro d’Ischia, ha investigat la biologia i l’ecologia d’una soca de cianobacteri formidial aïllada com a epífit de vida lliure damunt fulles de Posidonia oceanica. La identifiquen amb l’espècie Halomicronema metazoicum, cosa curiosa ja que les soques conegudes d’aquesta espècie són simbionts d’esponges marines. En aquest article, Mutalipassi et al. examinen el rang d’irradiació, de temperatura i de salinitat que permet el creixement d’aquest cianobacteri. Al mateix temps, examinen dins d’aquest rang la producció de compostos tòxics en un assaig in vivo emprat embrions de garota i rotífers adults. Han trobat, si més no, dos compostos bioactius, cadascun dels quals es produeix en diferents condicions d’irradiació i de temperatura. La major toxicitat del cianobacteri té lloc a temperatures i irradiacions elevades. Aquesta recerca obre, doncs, dos interrogants. D’una banda, caldrà aïllar i identificar els compostos tòxics, i les seves possibles aplicacions biotecnològiques. De l’altra, caldrà entendre quin és el paper ecològic que juguen aquests compostos tòxics en els prats de posidònies, més encara si atenem al fet que la tendència a l’augment de les temperatures de la Mar Mediterrània poden afectar els nivells d’aquestes i d’altres substàncies semblants.

Prat submarí de posidònies, fotografiat per Alberto Romeo en el 2007
De l’autoecologia d’un organisme a l’ecologia marina integrativa
Mirko Mutalipassi és investigador del Departament d’Ecologia Marina Integrativa del Centre d’Ecologia Bèntica de la Stazione Zoologica Anton Dohrn, a la Punta de San Pietro, a l’illa d’Ischia (al nord de la Badia de Nàpols). Mutalipassi ha realitzat els experiments en embrions d’eriçó de mar i en rotífers, així com les proves dels fotobioreactors, els cultius de cianobacteris i totes les fases de la recerca. Els treballs que es presenten en aquest article foren inclosos en la tesi doctoral que defensà reeixidament l’any passat a la UK Open University.
Valerio Mazzella fa la tesi doctoral sobre la resposta de les esponges marines a l’acidificació al mateix departament de Mutalipassi i Zupo. Mazzella col·laborà en aquesta recerca en la producció de cianobacteris en diferents condicions experimentals i ajudà en els bioassajos en embrions de garota i rotífers.
Giovanna Romano és investigadora del mateix Departament, però treballa des de la Villa Comunale en ecologia química i biotecnologia marina. Romano supervisà els experiments en embrions de garota.
Nadia Ruocco fa la tesi doctoral al Departament de Biologia i Evolució d’Organismes Marins (BEOM) de la Stazione Zoologica Anton Dohrn, a Villa Comunale, així com a la Unitat de Química Bio-Orgànica de l’Institut de Química Biomolecular de Pozzuoli i al Departament de Biologia de la Universitat de Nàpols.
Maria Costantini és investigadora del BEOM de Villa Comunale. Ruocco i Costantini realitzaren la identificació taxonòmica de les soques de cianobacteris.
Francesca Glaviano és investigadora del Departament d’Ecologia Marina Integrativa de la Punta San Pietro. En aquesta recerca col·laborà en les operacions del cultiu de cianobacteris.
Valerio Zupo, de Punta San Pietro, dissenyà el pla original de recerca i els fotobioreactors. Supervisà les activitats relacionades amb els cultius. La recerca fou finançada amb els ajuts de doctorat de Mutalipassi i Ruocco, i del projecte de recerca de Glaviano sobre l’ús sostenible de recursos marins.
Mutalipassi et al. agraeixen la col·laboració de la Unitat d’Anàlisi Morfo-funcional i Bioimatge (AMOBIO) de la mateixa Stazione Biologica, per les imatges de microscòpia electrònica fetes per F. Iamunno, R. Graziano i G. Lanzotti. També tenen un agraïment per a la Unitat de Monitoratge i Dades Ambientals (MEDA) per la recol·lecció de garotes, així com per a la Unitat de Recursos Marins per a la Recerca (MaRe, amb A. Macina i D. Caramiello) que d’aquestes garotes en preparà el gàmetes. Les fulles de posidònia emprades en aquesta recerca foren recollides pel vaixell de la Stazione, el Phoenicia del capità Rando.
El text de l’article fou redactat per Zupo, i revisat lingüísticament per R. Messina. Fou tramès a Biology Open el 3 d’abril. Després d’un procés de revisió fou acceptat el 7 d’octubre, i publicat el dia 15.
Aquesta recerca segueix una línia tradicional d’exploració de la diversitat de metabòlits secundaris dels cianobacteris. Per metabolisme secundari s’entén que no fa referència ni al metabolisme energètic (fotosíntesi, respiració, etc.) ni a la biosíntesi dels components majoritaris de l’organisme. Entre els metabòlits secundaris (carotenoids, ficobiliproteïnes, polisacàrids) podem trobar activitats amb aplicacions biotecnològiques: antibiòtics, conservants, protectors solars, etc. En les condicions adequades, soques de cianobacteris criades en fotobioreactors poden produir substàncies de gran interès.
Halomicronema metazoicum fou descrita originàriament com a cianobacteri simbiont de l’esponja Petrosia ficiformis. Cal dir que P. ficiformis té la capacitat de mantindre a ratlla depredadors com crustacis de la família dels artèmids o com eriçons de mar mitjançant toxines hemolítiques. Hom suposa que aquestes toxines són produïdes per cianobacteris simbionts. H. metazoicum pertany al grup B del complex de gèneres Lyngbya/Plectonema/Phormidium, que si bé no conformarien un grup natural sí tenen la característica comuna de produir exsudats tòxics.

‘Petrosia ficiformis’ és una esponja d’un color porpra característic que trobem en la Mediterrània i l’Atlàntic Nord
H. metazoicum forma filaments prims, consistents en cadenes de cèl·lules. Els treballs de Ruocco han mostrat la presència d’aquesta espècie com a epífit en fulles de Posidonia oceanica, herba que forma prades submarines. Un epífit no és ben bé un simbiont, ja que es limita a viure damunt de la planta. Però és evident que la producció de toxines per part de l’epífit pot protegir la posidònia de certs depredadors. Cal no oblidar tampoc que algunes de les toxines generades per explosions de creixement de cianobacteris poden ésser tòxiques per als humans a través de la xarxa tròfica.

Cianobacteris filamentosos observats al microscopi
El Phoenicia recollí les mostres de posidònia d’un part submarí que hi ha davant del litoral de la població de Lacco Ameno d’Ischia. Les mostres es recolliren durant la primavera. Si les posidònies formen prats damunt del fons sorrenc, cianobacteris formen tapissos damunt les fulles de la posidònia. Amb fòrceps esterilitzats, aquests tapissos foren recollits i transferits a plaques de multipou plenes de medi de cultiu. Els cultius eren renovats diverses vegades fins a assolir una completa purificació. Amb anàlisis morfològiques i proves de biologia molecular s’identificà entre els cultius una soca d’Halomicronema metazoicum. És la primera soca coneguda d’aquesta espècie que és de vida lliure, en el sentit que no és un simbiont d’esponges marines. Mutalipassi et al. sospiten que la facilitat amb la qual aquest cianobacteri creixia en un cultiu pur (axènic o unicianobacterià) s’explica per les propietats antibiòtics del seu propi exsudats. Sigui com sigui, cultivaren soques pures en recipients de vidre de 400 mL en una cambra termostatitzada a 18 °C, amb un nivell d’il·luminació de 200 μE, sota un cicle d’12 hores de fosca i 12 hores de llum. El medi era renovat cada 15 dies.
En aquests recipients, els cianobacteris tornaven a formar tapissos. Fragments d’aquests tapissos (de 5 grams de pes) eren subcultivats en Erlenmeyers amb 1,5 L de medi de cultiu. Els erlenmeyers eren dipositats en diferents condicions de llum, de temperatura i de salinitat, amb un total de 27 combinacions, i amb un duplicat per a cadascuna d’elles. Els nivells de salinitat (36%, 40%, 44%), de temperatura (16 °C, 18 °C, 22 °C) i d’irradiància (80, 140, 200 µE) foren definits d’acord amb el ventall que es troben a la Mar Mediterrània. Cada erlenmeyer era cultivat durant 40 dies, els suficients com per acumular productes del metabolisme secundari. Al final del període, es recollir el medi de cultiu, se’l filtrava (mida de porus de 0,22 μm) i se’l guardava en recipients de vidre conservats a -20 °C.
Dos organismes model per als estudis de toxicitat
Per estudiar la toxicitat dels exsudats cianobacterians es feien servir dos models biològics:
– adults del rotífer Brachionus plicatilis, procedents d’un cultiu continu mantingut a la Stazione Zoologica a 20 °C, alimentat a base d’un cultiu de l’alga verda Dunaliella.
– embrions de la garota Paracentrotus lividus.
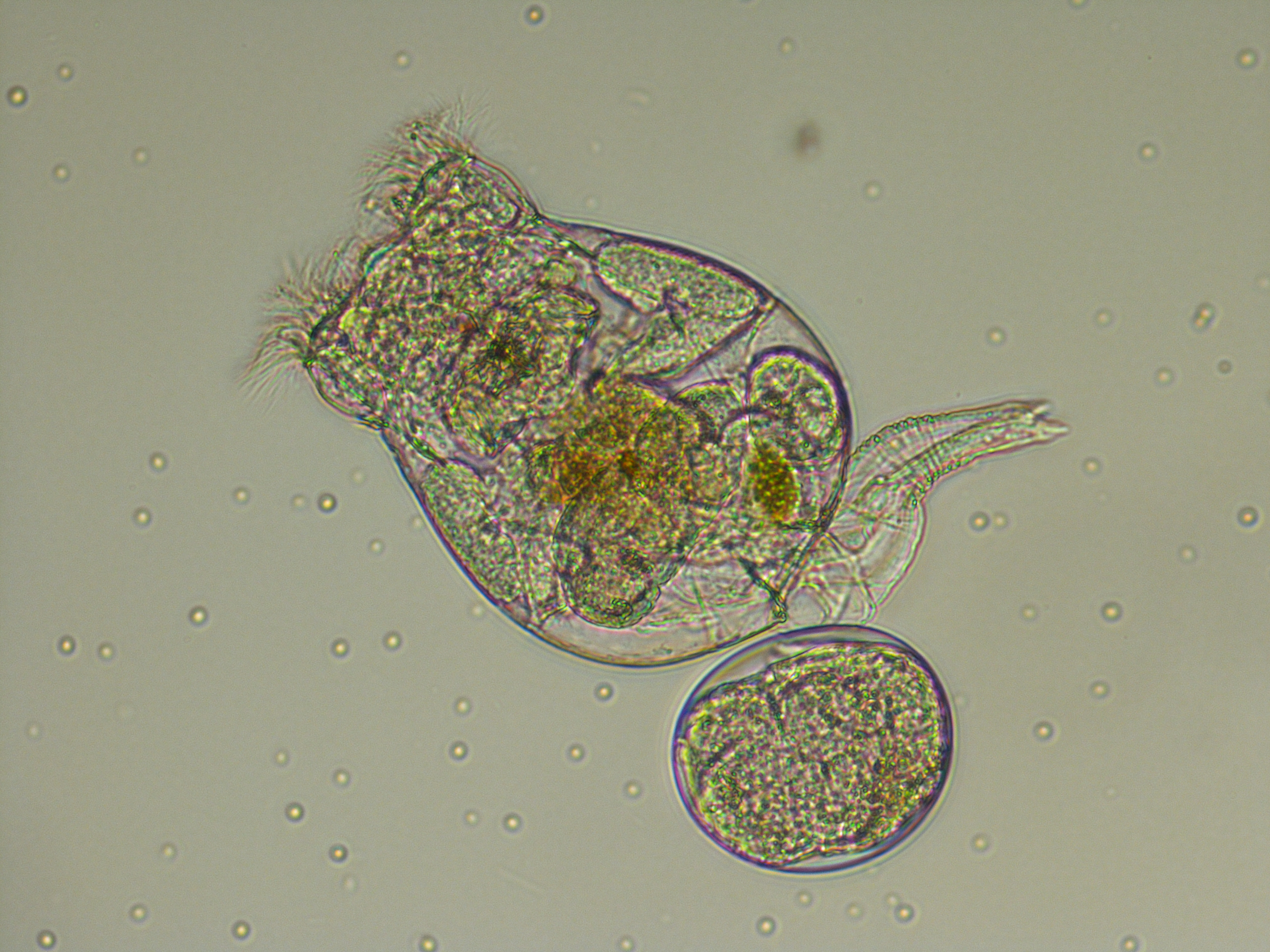
El cuc rotífer ‘Brachionus plicatilis’
Aquests organismes eren exposats a diferents concentracions dels medis de cultiu (1:5, 1:10, 1:100, 1:1000, 1:10000, 1:100000). Val a dir que les concentracions més elevades (1:5, 1:10, 1:100) produïen la mort immediata dels embrions de P. lividus. Contràriament, les concentracions més baixes (1:1000, 1:10000, 1:100000) eren del tot innòcues pels adults de B. plicatilis. Els dos models, doncs, reflecteixen dues condicions generals de susceptibilitat a toxines: si els embrions són molt sensibles, els rotífers adults disposen d’una maquinària potent de detoxificació.
En els experiments amb B. plicatilis, cada pou de placa de 5 mL contenia 50 individus adults. Hom examinava la motilitat i la taxa de supervivència cada 5 minuts (en la suspensió 1:5), cada 60 minuts (1:10) o cada 24 hores (1:100).
En els experiments amb embrions de P. lividus el punt de partida eren dues femelles madures i un mascle d’aquesta espècie. Als eriçons els injectaven 1 mL de KCl 0,5 M en el celoma a través de la boca per tal d’estimular la contracció de les gònades. Els sacsejaven vigorosament, i les femelles eren col·locades boca en amunt sobre un recipient de 50 mL. L’esperma era recollit en sec amb una pipeta Pasteur sobre la superfície dels gonòpors masculins. Els ous eren després fecundats amb aquest esperma. En cada experiment participaven un total de 500 embrions. El període experimental durava 48 hores, i es comptava el nombre de larves plutei normals que havien sobreviscut.
Els experiments amb aquests animals invertebrats es feren d’acord amb la directiva de la Unió Europea 2010/63/EU i la Declaració de Helsinki.
Creixement i toxicitat d’Halomicronema metazoicum
Les imatges de microscòpia electrònica mostren els tapissos d’H. metazoicum com integrats per filaments fins i homogenis de cèl·lules. Els filaments s’hi troben en una densitat considerable, lliures de contaminats, bo i que mostren petits agregats d’excusats mucosos. Després de 40 dies de cultiu, el medi és encara transparent, si bé amb una tonalitat brunosa.
En els experiments amb Brachionis plicatilis, la dada més informativa és la supervivència a les 24 hores. El nivell de toxicitat dels medis de cultiu depenia en bona mesura de la temperatura i de la irradiància. En el cas de la salinitat, les diferències es trobaven en les condicions més extremes de temperatura i d’irradiància. Les condicions de cultiu que condueixen a una major toxicitat són les de temperatura, salinitat i irradiància més elevades.
En els experiments amb embrions de garota, els patrons de toxicitat són més complexos, i varien d’acord amb el nivell de desenvolupament embrionari (segmentació, blastulació, gastrulació). La segmentació seria afectada per toxines constitutives, que varien poc d’acord amb la salinitat, la irradiància i la temperatura. En el cas de la gastrulació, la major toxicitat es troba en els medis de cianobacteris cultivats a temperatures i salinitats més altes. En el cas del desenvolupament cap a larves plutei, la major toxicitat es produeix en medis de cianobacteris cultivats a baixes temperatures.
Aquesta resposta bimodal, fa pensar Mutalipassi et al. que hi hauria, si més no, dos compostos bioactius, un que es produiria sobretot a temperatures de 16 °C (amb independència d’irradiància i de salinitat) i un altre a temperatures de 22 °C (que requeriria a més irradiàncies i salinitats intermèdies).
Una explicació d’aquestes diferències de toxicitat segons la temperatura es podria trobar en el fet que compostos tòxics de natura peptídica varien la seva bioactivitat segons com les condicions ambientals afecten la seva estructura.
El grup de Zupo treballa actualment en escatir la natura d’aquests compostos tòxics, així com el seu mecanisme d’acció.
Lligams:
– Growth and toxicity of Halomicronema metazoicum (Cyanoprokaryota, Cyanophyta) at different conditions of light, salinity and temperature. Mirko Mutalipassi, Valerio Mazzella, Giovanna Romano, Nadia Ruocco, Maria Costantini, Francesca Glaviano, Valerio Zupo Biol. Open bio.043604 (2019).
